The section of the Branchiopoda includes two groups differing even in their development,—the Phyllopoda and the Cladocera. The latter minute animals, provided with six pairs of foliaceous feet, which chiefly belong to the fresh waters, and are diffused under similar forms over the whole world, quit the egg with their full number of limbs. The Phyllopoda, on the contrary, in which the number of feet varies between 10 and 60 pairs, and some of which certainly live in the saturated lie of salterns and natron-lakes, but of which only one rather divergent genus (Nebalia) is found in the sea,1 have to undergo a metamorphosis. Mecznikow has recently observed the development of Nebalia, and concludes from his observations “that Nebalia, during its embryonal life, passes through the Nauplius- and Zoëa-stages, which in the Decapoda occur partly (in Penëus) in the free state.” “Therefore,” says he, “I regard Nebalia as a Phyllopodiform Decapod.” The youngest larvæ [of the Phyllopoda] are Nauplii, which we have already met with exceptionally in some Prawns, and which we shall now find reproduced almost without exception. The body-segments and feet, which are sometimes so numerous, are formed gradually from before backwards, without the indication of any sharply-discriminated regions of the body either by the time of their appearance or by their form. All the feet are essentially constructed in the same manner and resemble the maxillæ of the higher Crustacea.2 We might regard the Phyllopoda as Zoëæ which have not arrived at the formation of a peculiarly endowed abdomen or thorax, and instead of these have repeatedly reproduced the appendages which first follow the Nauplius-limbs.
Of the Copepoda—some of which, living in a free state, people the fresh waters, and in far more multifarious forms the sea, whilst others, as parasites, infest animals of the most various classes and often become wonderfully deformed—the developmental history, like their entire natural history, was, until lately, in a very unsatisfactory state. It is true, that we long ago knew that the Cyclopes of our fresh waters were excluded in the Nauplius-form, and that we were acquainted with some others of their young states; we had learnt, through Nordmann, that the same earliest form belonged to several parasitic Crustacea, which had previously passed, almost universally, as worms; but the connecting intermediate forms which would have permitted us to refer the regions of the body and the limbs of the larvæ to those of the adult animal, were wanting. The comprehensive and careful investigations of Claus have filled up this deficiency in our knowledge, and rendered the section of the Copepoda one of the best known in the whole class. The following statements are derived from the works of this able naturalist. From the abundance of valuable materials which they contain I select only those which are indispensable for the comprehension of the development of the Crustacea in general, because, in what relates to the Copepoda in particular, the facts have already been placed in the proper light by the representation of their most recent investigator, and must appear to any one whose eyes are open, as important evidence in favour of the Darwinian theory.3
All the larvæ of the free Copepoda investigated by Claus, have, at the earliest period, three pairs of limbs (the future antennæ and mandibles), the anterior with a single, and the two following ones with a double series of joints, or branchiæ. The unpaired eye, labrum, and mouth, already occupy their permanent positions. The posterior portion, which is usually short and destitute of limbs, bears two terminal setæ, between which the anus is situated. The form in this Nauplius-brood is extremely various,—it is sometimes compressed laterally, sometimes flat,—sometimes elongated, sometimes oval, sometimes round or even broader than long, and so forth. The changes which the first larval stages undergo during the progress of growth, consist essentially in an extension of the body and the sprouting forth of new limbs. “The following stage already displays a fourth pair of extremities, the future maxillæ.” Then follow at once three new pairs of limbs (the maxillipedes and the two anterior pairs of natatory feet). The larva still continues like a Nauplius, as the three anterior pairs of limbs represent rowing feet; at the next moult it is converted into the youngest Cyclops-like state, when it resembles the adult animal in the structure of the antennæ and buccal organs, although the number of limbs and body segments is still much less, for only the rudiments of the third and fourth pairs of natatory feet have made their appearance in the form of cushions fringed with setæ, and the body consists of the oval cephalothorax, the second, third, and fourth thoracic segments, and an elongated terminal joint. In the Cyclopidæ the posterior antennæ have lost their secondary branch, and the mandibles have completely thrown off the previously existing natatory feet, whilst in the other families these appendages persist, more or less altered. “Beyond this stage of free development, many forms of the parasitic Copepoda, such as Lernanthropus and Chondracanthus, do not pass, as they do not acquire the third and fourth pairs of limbs, nor does a separation of the fifth thoracic segment from the abdomen take place; others (Achtheres) even fall to a lower grade by the subsequent loss of the two pairs of natatory feet. But all free Copepoda, and most of the parasitic Crustacea, pass through a longer or shorter series of stages of development, in which the limbs acquire a higher degree of division into joints in continuous sequence, the posterior pairs of feet are developed, and the last thoracic segment and the different abdominal segments are successively separated from the common terminal portion.”
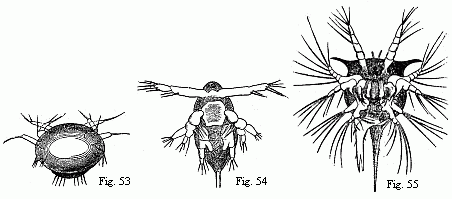
Fig. 53 and 54. Nauplii of Copepoda, the former magnified, the latter magnified 2x.
Fig. 55. Nauplius of Tetraclita porosa after the first moult, magnified. The brain is seen surrounding the eye, and from it the olfactory filaments issue; behind it are some delicate muscles passing to the buccal hood.
There is only one thing more to be indicated in the developmental history of the parasitic Crustacea, namely that some of them, such as Achtheres percarum, certainly quit the egg like the rest in a Nauplius-like form, inasmuch as the plump, oval, astomatous body bears two pairs of simple rowing feet, and behind these, as traces of the third pair, two inflations furnished each with a long seta, but that beneath this Nauplius-skin a very different larva lies ready prepared, which in a few hours bursts its clumsy envelope and then makes its appearance in a form “which agrees in the segmentation of the body and in the development of the extremities with the first Cyclops-stage” (Claus). The entire series of Nauplius-stages which are passed through by the free Copepoda, are in this case completely over-leapt.
A final and very peculiar section of the Crustacea is formed by the two orders of the Cirripedia and Rhizocephala.4
In these also the brood bursts out in the Nauplius-form, and speedily strips off its earliest larva-skin which is distinguished by no peculiarities worth noticing. Here also we find again the same pyriform shape of the unsegmented body, the same number and structure of the feet, the same position of the median eye (which, however, is wanting in Sacculina purpurea, and according to Darwin in some species of Lepas), and the same position of the “buccal hood,” as in the Nauplii of the Prawns and Copepoda. From the latter the Nauplii of the Cirripedia and Rhizocephala are distinguished by the possession of a dorsal shield or carapace, which sometimes (Sacculina purpurea) projects far beyond the body all round; and they are distinguished not only from other Nauplii, but as far as I know from all other Crustacea, by the circumstance that structures which are elsewhere combined with the two anterior limbs (antennæ), here occur separated from them.
The anterior antennæ of the Copepoda, Cladocera, Phyllopoda (Leydig, Claus), Ostracoda (at least the Cypridinæ), Diastylidæ, Edriophthalma, and Podophthalma, with few exceptions relating to terrestrial animals or parasites, bear peculiar filaments which I have already repeatedly mentioned as “olfactory filaments.” A pair of similar filaments spring, in the larvæ of the Cirripedia and Rhizocephala, directly from the brain.
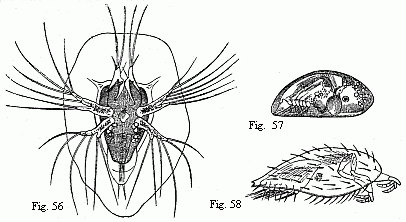
Fig. 56. Nauplius of Sacculina purpurea, shortly before the second moult, magnified. We may recognise in the first pair of feet the future adherent feet, and in the abdomen six pairs of natatory feet with long setæ.
Fig. 57. Pupa of a Balanide (Chthamalus ?), magnified. The adherent feet are retracted within the rather opaque anterior part of the shell.
Fig. 58. Pupa of Sacculina purpurea, magnified. The filaments on the adherent feet may be the commencements of the future roots.
At the base of the inferior antennæ in the Decapoda the so-called “green-gland” has its opening; in the Macrura at the end of a conical process. A similar conical process with an efferent duct traversing it is very striking in most of the Amphipoda. In the Ostracoda, Zenker describes a gland situated in the base of the inferior antennæ, and opening at the extremity of an extraordinarily long “spine.” In the Nauplii of Cyclops and Cyclopsine, Claus finds pale “shell-glands,” which commence in the intermediate pair of limbs (the posterior antennæ). On the other hand in the Nauplii of the Cirripedia and Rhizocephala the “shell-glands” open at the ends of conical processes, sometimes of most remarkable length, which spring from the angles of the broad frontal margin, and have been interpreted sometimes as antennæ (Burmeister, Darwin) and sometimes as mere “horns of the carapace” (Krohn). The connexion of the “shell-glands” with the frontal horns has been recognised unmistakably in the larvæ of Lepas, and indeed the resemblance of the frontal horns with the conical processes on the inferior antennæ of the Amphipoda, is complete throughout.5
Notwithstanding their agreement in this important peculiarity, the Nauplii of these two orders present material differences in many other particulars. The abdomen of the young Cirripede is produced beneath the anus into a long tail-like appendage which is furcate at the extremity, and over the anus there is a second long, spine-like process; the abdomen in the Rhizocephala terminates in two short points,—in a “moveable caudal fork, as in the Rotatoria,” (O. Schmidt). The young Cirripedes have a mouth, stomach, intestine, and anus, and their two posterior pairs of limbs are beset with multifarious teeth, setæ, and hooks, which certainly assist in the inception of nourishment. All this is wanting in the young Rhizocephala. The Nauplii of the Cirripedia have to undergo several moults whilst in that form; the Nauplii of the Rhizocephala, being astomatous, cannot of course live long as Nauplii, and in the course of only a few days they become transformed into equally astomatous “pupæ,” as Darwin calls them.
The carapace folds itself together, so that the little animal acquires the aspect of a bivalve shell, the foremost limbs become transformed into very peculiar adherent feet (“prehensile antennæ,” Darwin), and the two following pairs are cast off; like the frontal horns. On the abdomen six pairs of powerful biramose natatory feet with long setæ have been formed beneath the Nauplius-skin, and behind these are two short, setigerous caudal appendages (Fig. 58).
The pupæ of the Cirripedia (Fig. 57), which are likewise astomatous, agree completely in all these parts with those of the Rhizocephala, even to the minutest details of the segmentation and bristling of the natatory feet;6 they are especially distinguished from them by the possession of a pair of composite eyes. Sometimes also traces of the frontal horns seem to persist.7
As the Cirripedia and Rhizocephala now in general resemble each other far more than in their Nauplius-state, this is also the case with the individual members of each of the two orders.
The pupæ in both orders attach themselves by means of the adherent feet; those of the Cirripedes to rocks, shells, turtles, drift-wood, ships, etc.,—those of the Rhizocephala to the abdomen of Crabs, Porcellanæ, and Hermit Crabs. The carapace of the Cirripedes becomes converted, as is well-known, into a peculiar test, on account of which they were formerly placed among the Mollusca, and the natatory feet grow into long cirri, which whirl nourishment towards the mouth, which is now open. The Rhizocephala remain astomatous; they lose all their limbs completely, and appear as sausage-like, sack-shaped or discoidal excrescences of their host, filled with ova (Figs. 59, 60); from the point of attachment closed tubes, ramified like roots, sink into the interior of the host, twisting round its intestine, or becoming diffused among the sac-like tubes of its liver. The only manifestations of life which persist in these non plus ultras in the series of retrogressively metamorphosed Crustacea, are powerful contractions of the roots, and an alternate expansion and contraction of the body, in consequence of which water flows into the brood-cavity and is again expelled, through a wide orifice.8
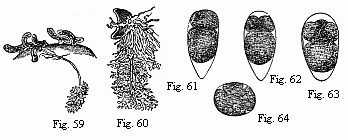
Fig. 59. Young of Peltogaster socialis on the abdomen of a small Hermit Crab; in one of them the fasciculately ramified roots in the liver of the Crab are shown. Animal and roots deep yellow.
Fig. 60. Young Sacculina purpurea with its roots; the animal purple-red, the roots dark grass-green. Magnified.
Figs. 61–63. Eggs of Tetraclita porosa in segmentation, magnified. The larger of the two first-formed spheres of segmentation is always turned towards the pointed end of the egg.
Fig. 64. Egg of Lernæodiscus Porcellanæ, in segmentation, magnified.
Out of several Cirripedes, which are anomalous both in structure and development, Cryptophialus minutus must be mentioned here; Darwin found it in great quantities together in the shell of Concholepas peruviana on the Chonos Islands. The egg, which is at first elliptical, soon, according to Darwin, becomes broader at the anterior extremity, and acquires three club-shaped horns, one at each anterior angle and one behind; no internal parts can as yet be detected. Subsequently the posterior horn disappears, and the adherent feet may be recognised within the anterior ones. From this “egg-like larva”—(Darwin says of it, “I hardly know what to call it”)—the pupa is directly produced. Its carapace is but slightly compressed laterally and hairy, as in Sacculina purpurea; the adherent feet are of considerable size, and the natatory feet are wanting, as, in the adult animal, are the corresponding cirri. As I learn from Mr. Spence Bate, the Nauplius-stage appears to be overleaped and the larvæ to leave the egg in the pupa-form, in the case of a Rhizocephalon (Peltogaster ?) found by Dr. Powell in the Mauritius.
I will conclude this general view with a few words upon the earliest processes in the development of the Crustacea. Until recently it was regarded as a general rule that, by the partial segmentation of the vitellus a germinal disc was formed, and in this, corresponding to the ventral surface of the embryo, a primitive band. We now know that in the Copepoda (Claus), in the Rhizocephala (Fig. 64), and, as I can add, in the Cirripedia (Figs. 61–63) the segmentation is complete, and the embryos are sketched out in their complete form without any preceding primitive band. Probably the latter will always be the case where the young are hatched as true Nauplii (and not merely with a Nauplius-skin, as in Achtheres). The two modes of development may occur in very closely allied animals, as is proved by Achtheres among the Copepoda.9
1. If the Phyllopoda may be regarded as the nearest allies of the Trilobites, they would furnish, with Lepidosteus and Polypterus, Lepidosiren and Protopterus, a further example of the preservation in fresh waters of forms long since extinguished in the sea. The occurrence of the Artemiæ in supersaline water would at the same time show that they do not escape destruction by means of the fresh water, but in consequence of the less amount of competition in it.
2. “The maxilla of the Decapod-larva (Krebslarve) is a sort of Phyllopodal foot” (Claus).
3. I am still unacquainted with Claus’ latest and larger work, but no doubt the same may be said of it.
4. The most various opinions prevail as to the position of the Cirripedia. Some ascribe to them a very subordinate position among the Copepoda; as Milne-Edwards (1852). In direct opposition to this notion of his father’s, Alph. Milne-Edwards places them (as Basinotes) opposite to all the other Crustacea (Eleuthéronotes). Darwin regards them as forming a peculiar sub-class equivalent to the Podophthalma, Edriophthalma, etc. This appears to me to be most convenient. I would not combine the Rhizocephala with the Cirripedia, as Liljeborg has done, but place them in opposition as equivalent, like the Amphipoda and Isopoda. The near relationship of the Cirripedia to the Ostracoda is also spoken of, but the similarity of the so-called “Cypris-like larvæ,” or Cirriped-pupæ as Darwin calls them, to Cypris is so purely external, even as regards the shell, that the relationship appears to me to be scarcely greater than that of Peltogaster socialis (Fig. 59) with the family of the sausages.
5. In connexion with this it may be mentioned that, in the females of Brachyscelus, in which the posterior antennæ are deficient, the conical processes with the canal permeating them are nevertheless retained.
6. Compare the figure given by Darwin (Balanidæ Pl. xxx fig. 5) of the first natatory foot of the pupa of Lepas australis, with that of Lernæodiscus Porcellanæ published in the ‘Archiv für Naturgeschichte’ (1863, Taf. iii, fig. 5). The sole distinction, that in the latter there are only 3 setæ at the end of the outer branch, whilst in the Cirripedia there are 4 on the first and 5 on the following natatory feet, may be due to an error on my part.
7. Darwin describes as “acoustic orifices” small apertures in the shell of the pupæ of the Cirripedia, which, frequently surrounded by a border, are situated, in Lepas pectinata, upon short, horn-like processes. I feel scarcely any hesitation in regarding the apertures as those of the “shell-glands,” and the horn-like processes as remains of the frontal horns.
8. The roots of Sacculina purpurea (Fig. 60) which is parasitic upon a small Hermit Crab, are made use of by two parasitic Isopods, namely a Bopyrus and the before mentioned Cryptoniscus planarioides (Fig. 42). These take up their abode beneath the Sacculina and cause it to die away by intercepting the nourishment conveyed by the roots; the roots, however, continue to grow, even without the Sacculina, and frequently attain an extraordinary extension, especially when a Bopyrus obtains its nourishment from them.
9. I have not mentioned the Pycnogonidæ, because I do not regard them as Crustacea; nor the Xiphosura and Trilobites, because, having never investigated them myself, I knew too little about them, and especially because I am unacquainted with the details of the explanations given by Barrande of the development of the latter. According to Mr. Spence Bate “the young of Trilobites are of the Nauplius-form.”)
{kind=link}