
Less varied than that of the Stalk-eyed Crustacea is the mode of development of the Isopoda and Amphipoda, which Leach united in the section Edriophthalma, or Crustacea with sessile eyes.
The Rock-Slaters (Ligia) may serve as an example of the development of the Isopoda. In these, as in Mysis, the caudal portion of the embryo is bent not downwards, but upwards; as in Mysis also, a larval membrane is first of all formed, within which the Slater is developed. In Mysis this first larval skin may be compared to a Nauplius; in Ligia it appears like a maggot quite destitute of appendages, but produced into a long simple tail (Fig. 37). The egg-membrane is retained longer than in Mysis; it bursts only when the limbs of the young Slater are already partially developed in their full number. The dorsal surface of the Slater is united to the larval skin a little behind the head. At this point, when the union has been dissolved a little before the change of skin, there is a foliaceous appendage, which exists only for a short time, and disappears before the young Slater quits the brood-pouch of the mother.
Fig. 36. Embryo of Ligia in the egg,
magnified. D. yelk; L. liver.
Fig. 37. Maggot-like larva of Ligia, magnified. R
remains of the egg-membrane. We see on the lower surface, from
before backwards:—the anterior and posterior antennæ,
the mandibles, the anterior and posterior maxillæ,
maxillipedes, six ambulatory feet, the last segment of the
middle-body destitute of appendages, five abdominal feet, and the
caudal feet.
The young animal, when it begins to take care of itself, resembles the old ones in almost all parts, except one important difference; it possesses only six, instead of seven pairs of ambulatory feet; and the last segment of the middle-body is but slightly developed and destitute of appendages. It need hardly be mentioned that the sexual peculiarities are not yet developed, and that in the males the hand-like enlargements of the anterior ambulatory feet and the copulatory appendages are still deficient.
To the question, how far the development of Ligia is repeated in the other Isopoda, I can only give an unsatisfactory answer. The curvature of the embryo upwards instead of downwards was met with by me as well as by Rathke in Idothea, and likewise in Cassidina, Philoscia, Tanais, and the Bopyridæ,—indeed, I failed to find it in none of the Isopoda examined for this purpose. In Cassidina also the first larval skin without appendages is easily detected; it is destitute of the long tail, but is strongly bent in the egg, as in Ligia, and consequently cannot be mistaken for an “inner egg-membrane.” This, however, might happen in Philoscia, in which the larval skin is closely applied to the egg-membrane (Fig. 38), and is only to be explained as the larval skin by a reference to Ligia and Cassidina. The foliaceous appendage on the back has long been known in the young of the common Water Slater (Asellus).1 That the last pair of feet of the thorax is wanting in the young of the Wood-lice (Porcellionides, M.-Edw.) and Fish-lice (Cymothoadiens, M.-Edw.) has already been noticed by Milne-Edwards. This applies also to the Box-Slaters (Idothea), to the viviparous Globe-Slaters (Sphæroma) and Shield-Slaters (Cassidina), to the Bopyridæ (Bopyrus, Entoniscus, Cryptoniscus, n.g.), and to the Cheliferous Slaters (Tanais), and therefore probably to the great majority of the Isopoda. All the other limbs are usually well developed in the young Isopoda. In Tanais alone, all the abdominal feet are wanting (but not those of the tail); they are developed simultaneously with the last pair of feet of the thorax.

Fig. 38. Embryo of a Philoscia in the egg,
magnified.
Fig. 39. Embryo of Cryptoniscus planarioides, magnified.
Fig. 40. Last foot of the middle-body of the larva of Entoniscus
Porcellanæ, magnified.
The last pair of feet on the middle-body of the larva, consequently the penultimate pair in the adult animal, is almost always similar in structure to the preceding pair. A remarkable exception is, however, presented in this respect by Cryptoniscus and Entoniscus,—remarkable as a confirmation of Darwin’s proposition that “parts developed in an unusual manner are very variable,” for in the peculiarly-formed pair of feet there exists the greatest possible difference between the three species hitherto observed. In Cryptoniscus (Fig. 39) this last foot is thin and rod-like; in Entoniscus Cancrorum remarkably long and furnished with a strongly thickened hand and a peculiarly constructed chela; in Entoniscus Porcellanæ very short, imperfectly jointed, and with a large ovate terminal joint (Fig. 40).
Some Isopods undergo a considerable change immediately before the attainment of sexual maturity. This is the case with the males of Tanais which have already been noticed, and, according to Hesse, with the Pranizæ, in which both sexes are said to pass into the form known as Anceus. But Spence Bate, a careful observer, states that he has seen females of the form of Praniza laden with eggs far advanced in their development.
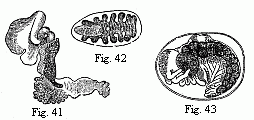
Fig. 41. Entoniscus Cancrorum, female,
magnified.
Fig. 42. Cryptoniscus planarioides, female, magnified.
Fig. 43. Embryo of a Corophium, magnified.
In this order we meet for the first time with an extensive retrograde metamorphosis as a consequence of a parasitic mode of life. Even in some Fish-lice (Cymothoa) the young are lively swimmers, and the adults stiff, stupid, heavy fellows, whose short clinging feet are capable of but little movement. In the Bopyridæ (Bopyrus, Phryxus, Kepone, etc., which might have been conveniently left in a single genus), which are parasitic on Crabs, Lobsters, etc., taking up their abode chiefly in the branchial cavity, the adult females are usually quite destitute of eyes; the antennæ are rudimentary; the broad body is frequently unsymmetrically developed in consequence of the confined space; its segments are more or less amalgamated with each other; the feet are stunted, and the appendages of the abdomen transformed from natatory feet with long setæ into foliaceous or tongue-shaped and sometimes ramified branchiæ. In the dwarfish males the eyes, antennæ, and feet, are usually better preserved than in the females; but on the other hand all the appendages of the abdomen have not unfrequently disappeared, and sometimes every trace of segmentation. In the females of Entoniscus, which are found in the body-cavity of Crabs and Porcellanæ, the eyes, antennæ, and buccal organs, the segmentation of the vermiform body, and in one species (Fig. 41) the whole of the limbs, disappear almost without leaving a trace; and Cryptoniscus planarioides would almost be regarded as a Flatworm rather than an Isopod, if its eggs and young did not betray its Crustacean nature. Among the males of these various Bopyridæ, that of Entoniscus Porcellanæ occupies the lowest place; it is confined all its life to six pairs of feet, which are reduced to shapeless rounded lumps.
The Amphipoda are distinguishable from the Isopoda at an early period in the egg by the different position of the embryo, the hinder extremity of which is bent downwards. In all the animals of this order which have been examined for it,2 a peculiar structure makes its appearance very early on the anterior part of the back, by which the embryo is attached to the “inner egg-membrane,” and which has been called the “micropylar apparatus,” but improperly as it seems to me.3 It will remind us of the union of the young Isopoda with the larval membrane and of the unpaired “adherent organ” on the nape of the Cladocera, which is remarkably developed in Evadne and persists throughout life; but in Daphnia pulex, according to Leydig, although present in the young animals, disappears without leaving a trace in the adults.
The young animal, whilst still in the egg, acquires the full number of its segments and limbs. In cases where segments are amalgamated together, such as the last two segments of the thorax in Dulichia, the last abdominal segments and the tail in Gammarus ambulans and Corophium dentatum, n. sp., and the last abdominal segments and the tail in Brachyscelus,4 or where one or more segments are deficient, as in Dulichia and the Caprellæ, we find the same fusion and the same deficiencies in young animals taken out of the brood-pouch of their mother. Even peculiarities in the structure of the limbs, so far as they are common to both sexes, are usually well-marked in the newly hatched young, so that the latter generally differ from their parents only by their stouter form, the smaller number of the antennal joints and olfactory filaments, and also of the setæ and teeth with which the body or feet are armed, and perhaps by the comparatively larger size of the secondary flagellum. An exception to this rule is presented by the Hyperinæ which usually live upon Acalephæ. In these the young and adults often have a remarkably different appearance; but even in these there is no new formation of body-segments and limbs, but only a gradual transformation of these parts.5
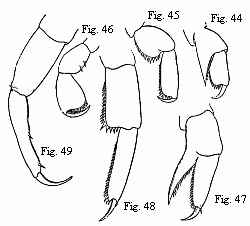
Figs. 44–46. Feet of a half-grown Hyperia
Martinezii, n. sp. (Named after my valued friend the amiable
Spanish zoologist, M. Francisco de Paula Martinez y Saes, at
present on a voyage round the world.)
Figs. 47–49. Feet of a nearly adult male of the same species;
44 and 47 from the first pair of anterior feet (gnathopoda); 44 and
48 from the first, and 46 and 49 from the last pair of thoracic
feet. Magnified.
Thus, in order to give a few examples, the powerful chelæ of the antepenultimate pair of feet, of Phromina sedentaria, are produced, according to Pagenstecher, from simple feet of ordinary structure; and vice versà, the chelæ on the penultimate pair of feet of the young Brachyscelus, become converted into simple feet. In the young of the last-mentioned genus the long head is drawn out into a conical point and bears remarkably small eyes; in course of growth, the latter, as in most of the Hyperinæ, attain an enormous size, and almost entirely occupy the head, which then appears spherical, etc.
The difference of the sexes which, in the Gammarinæ is usually expressed chiefly in the structure of the anterior feet (gnathopoda, Sp. Bate) and in the Hyperinæ in the structure of the antennæ, is often so great that males and females have been described as distinct species, and even repeatedly placed in different genera (Orchestia and Talitrus, Cerapus and Dercothoë, Lestrigonus and Hyperia) or even families (Hypérines anormales and Hypérines ordinaires). Nevertheless it is only developed when the animals are nearly full-grown. Up to this period the young resemble the females in a general way, even in some cases in which these differ more widely than the males from the “Type” of the order. Thus in the male Shore-hoppers (Orchestia) the second pair of the anterior feet is provided with a powerful hand, as in the majority of the Amphipoda, but very differently constructed in the females. The young, nevertheless, resemble the female. Thus also,—and this is an extremely rare case,6—the females of Brachyscelus are destitute of the posterior (or inferior) antennæ; the male possesses them like other Amphipodæ; in the young I, like Spence Bate, can find no trace of them.
It is, however, to be particularly remarked, that the development of the sexual peculiarities does not stand still on the attainment of sexual maturity.
For example, the younger sexually mature males of Orchestia Tucurauna, n. sp., have slender inferior antennæ, with the joints of the flagellum not fused together, the clasping margin (“palm,” Sp. Bate) of the hand in the second pair of feet is uniformly convex, the last pair of feet is slender and similar to the preceding. Subsequently the antennæ become thickened, two, three, or four of the first joints of the flagellum are fused together, the palm of the hand acquires a deep emargination near its inferior angle, and the intermediate joints of the last pair of feet become swelled into a considerable incrassation. No museum-zoologist would hesitate about fabricating two distinct species, if the oldest and youngest sexually mature males were sent to him without the uniting intermediate forms. In the younger males of Orchestia Tucuratinga, although the microscopic examination of their testes showed that they were already sexually mature, the emargination of the clasping margin of the hand (represented in Fig. 50) and the corresponding process of the finger, are still entirely wanting. The same may be observed in Cerapus and Caprella, and probably in all cases where hereditary sexual differences occur.
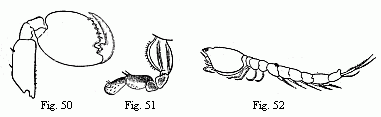
Fig. 50. Foot of the second pair (“second
pair of gnathopoda”) of the male of Orchestia
Tucurauna, magnified.
Fig. 51. Foot of the second pair (“second pair of
gnathopoda”) of the female of Orchestia Tucurauna,
magnified.
Fig. 52. Male of a Bodotria, magnified. Note the long
inferior antennæ, which are closely applied to the body, and
of which the apex is visible beneath the caudal appendages.
Next to the extensive sections of the Stalk-eyed and Sessile-eyed Crustacea, but more nearly allied to the former than to the latter, comes the remarkable family of the Diastylidæ or Cumacea. The young, which Kröyer took out of the brood-pouch of the female, and which attained one-fourth of the length of their mother, resembled the adult animals almost in all parts. Whether, as in Mysis and Ligia, a transformation occurs within the brood-pouch, which is constructed in the same way as in Mysis, is not known.7 The caudal portion of the embryo in the Diastylidæ, as I have recently observed, is curved upwards as in the Isopoda, and the last pair of feet of the thorax is wanting.
Equally scanty is our knowledge of the developmental history of the Ostracoda. We know scarcely anything except that their anterior limbs are developed before the posterior one (Zenker). The development of Cypris has recently been observed by Claus:—“The youngest stages are shell-bearing Nauplius-forms.”
1. Leydig has compared this
foliaceous appendage of the Water Slaters with the “green
gland” or “shell-gland” of other crustacea,
assuming that the green gland has no efferent duct and appealing to
the fact that the two organs occur “in the same place.”
This interpretation is by no means a happy one. In the first place
we may easily ascertain in Leucifer, as was also found to be
the case by Claus, that the “green gland” really opens
at the end of the process described by Milne-Edwards as a
“tubercule auditif” and by Spence Bate as an
“olfactory denticle.” And, secondly, the position is
about as different as it can well be. In the one case a paired
gland, opening at the base of the posterior antennæ, and
therefore on the lower surface of the second segment; in the
other an unpaired structure rising in the median line of the back
behind the seventh segment, (“behind the boundary line
of the first thoracic segment,” Leydig).
2. In the genera Orchestoidea, Orchestia,
Allorchestes, Montagua, Batea n.g., Amphilochus, Atylus,
Microdeutopus, Leucothoë, Melita, Gammarus (according to
Meissner and La Valette), Amphithoë, Cerapus, Cyrtophium,
Corophium, Dulichia, Protella and Caprella.
3. Little as a name may actually affect the facts,
we ought certainly to confine the name “micropyle” to
canals of the egg-membrane, which serve for the entrance of the
semen. But the outer egg-membrane passes over the “micropylar
apparatus” of the Amphipoda without any perforation,
according to Meissner’s and La Valette’s own
statements; it appears never to be present before fecundation,
attains its greatest development at a subsequent period of the
ovular life, and the delicate canals which penetrate it do not even
seem to be always present, indeed it seems to belong to the embryo
rather than to the egg-membrane. I have never been able to convince
myself that the so-called “inner egg-membrane” is
really of this nature, and not perhaps the earliest larva skin, not
formed until after impregnation, as might be supposed with
reference to Ligia, Cassidina and Philoscia.
4. According to Spence Bate, in Brachyscelus
crusculum the fifth abdominal segment is not amalgamated with
the sixth (the tail) but with the fourth, which I should be
inclined to doubt, considering the close agreement which this
species otherwise shows with the two species that I have
investigated.
5. In the young of Hyperia galba Spence
Bate did not find any of the abdominal feet, or the last two pairs
of thoracic feet, but this very remarkable statement required
confirmation the more because he examined these minute animals only
in the dried state. Subsequently I had the wished-for opportunity
of tracing the development of a Hyperia which is not
uncommon upon Ctenophora, especially Beroë gilva,
Eschsch. The youngest larva from the brood-pouch of the mother
already possess the whole of the thoracic feet; on the other
hand, like Spence Bate, I cannot find those of the abdomen. At
first simple enough, all these feet soon become converted, like the
anterior feet, into richly denticulated prehensile feet, and indeed
of three different forms, the anterior feet (Fig. 44) the two
following pairs (Fig. 45) and finally the three last pairs (Fig.
46) being similarly constructed and different from the rest. In
this form the feet remain for a very long time, whilst the
abdominal appendages grow into powerful natatory organs, and the
eyes, which at first seemed to me to be wanting, into large
hemispheres. In the transition to the form of the adult animal the
last three pairs of feet (Fig. 49) especially undergo a
considerable change. The difference between the two sexes is
considerable; the females are distinguished by a very broad thorax,
and the males (Lestrigonus) by very long antennæ, of
which the anterior bear an unusual abundance of olfactory
filaments.
Their youngest larvæ of course cannot swim;
they are helpless little animals which firmly cling especially to
the swimming laminæ of their host; the adult
Hyperiæ, which are not unfrequently met with free in the
sea, are, as is well known, the most admirable swimmers in their
order. (“Il nage avec une rapidité
extrême,” says Van Beneden of H. Latreillii
M.-Edw.)
The transformation of the Hyperiæ is
evidently to be regarded as acquired and not
inherited, that is to say the late appearance of the abdominal
appendages and the peculiar structure of the feet in the young are
not to be brought into unison with the historical development of
the Amphipoda, but to be placed to the account of the parasitic
mode of life of the young.
As in Brachyscelus, free locomotion has
been continued to the adult and not to the young, contrary to the
usual method among parasites. Still more remarkable is a similar
circumstance in Caligus, among the parasitic Copepoda. The
young animal, described by Burmeister as a peculiar genus,
Chalimus, lies at anchor upon a fish by means of a cable
springing from its forehead, and having its extremity firmly seated
in the skin of the fish. When sexual maturity is attained, the
cable is cut, and the adult Caligi, which are admirable
swimmers, are not unfrequently captured swimming freely in the sea.
(See ‘Archiv. für Naturgeschichte’ 1852, I. p.
91).
6. “I know of no case in which the inferior
(antennæ) are obsolete, when the superior are
developed,” Dana. (Darwin, ‘Monograph on the Subclass
Cirripedia, Lepadidæ’ p. 15.)
7. A trustworthy English Naturalist, Goodsir,
described the brood-pouch and eggs of Cuma as early as 1843.
Kröyer, whose painstaking care and conscientiousness is
recognised with wonder by every one who has met him on a common
field of work, confirmed Goodsir’s statements in 1846, and,
as above mentioned, took out of the brood-pouch embryos advanced in
development and resembling their parents. By this the question
whether the Diastylidæ are full-grown animals or larvæ,
is completely and for ever set at rest, and only the famous names
of Agassiz, Dana and Milne-Edwards, who would recently reduce them
again to larvæ (see Van Beneden, ‘Rech. sur la Fauna
littor. de Belgique’ Crustacées, pp. 73, 74), induce
me, on the basis of numerous investigations of my own, to declare
in Van Beneden’s words; “Parmi toutes les formes
embryonnaires de podophthalmes ou d’édriophthalmes que
nous avons observées sur nos côtes, nous n’en
avons pas vu une seule qui eût même la moindre
resemblance avec un Cuma quelconque.” The only
thing that suits the larvæ of Hippolyte,
Palæmon and Alpheus, in the family character of
the Cumacea as given by Kröyer which occupies three pages
(Kröyer, ‘Naturh. Tidsskrift, Ny Raekke,’ Bd. ii.
pp. 203–206) is: “Duo antennarum paria.” And
this, as is well known, applies to nearly all Crustacea. How well
warranted are we therefore in identifying the latter with the
former. However, it is sufficient for any one to glance at the
larva of Palæmon (Fig. 27) and the Cumacean (Fig. 52) in
order to be convinced of their extraordinary similarity!