THE EVOLUTION OF MAN
Volume II
CHAPTER XXVII
THE EVOLUTION OF THE ALIMENTARY SYSTEM
The chief of the vegetal organs of the human frame, to the evolution of which we now turn our attention, is the alimentary canal. The gut is the oldest of all the organs of the metazoic body, and it leads us back to the earliest age of the formation of organs—to the first section of the Laurentian period. As we have already seen, the result of the first division of labour among the homogeneous cells of the earliest multicellular animal body was the formation of an alimentary cavity. The first duty and first need of every organism is self-preservation. This is met by the functions of the nutrition and the covering of the body. When, therefore, in the primitive globular Blastæa the homogeneous cells began to effect a division of labour, they had first to meet this twofold need. One half were converted into alimentary cells and enclosed a digestive cavity, the gut. The other half became covering cells, and formed an envelope round the alimentary tube and the whole body. Thus arose the primary germinal layers—the inner, alimentary, or vegetal layer, and the outer, covering, or animal layer. (Cf. pp. 214–17.)
When we try to construct an animal frame of the simplest conceivable type, that has some such primitive alimentary canal and the two primary layers constituting its wall, we inevitably come to the very remarkable embryonic form of the gastrula, which we have found with extraordinary persistence throughout the whole range of animals, with the exception of the unicellulars—in the Sponges, Cnidaria, Platodes, Vermalia, Molluscs, Articulates, Echinoderms, Tunicates, and Vertebrates. In all these stems the gastrula recurs in the same very simple form. It is certainly a remarkable fact that the gastrula is found in various animals as a larva-stage in their individual development, and that this gastrula, though much disguised by cenogenetic modifications, has everywhere essentially the same palingenetic structure (Figs. 30–35). The elaborate alimentary canal of the higher animals develops ontogenetically from the same simple primitive gut of the gastrula.
This gastræa theory is now accepted by nearly all zoologists. It was first supported and partly modified by Professor Ray-Lankester; he proposed three years afterwards (in his essay on the development of the Molluscs, 1875) to give the name of archenteron to the primitive gut and blastoporus to the primitive mouth.
Before we follow the development of the human alimentary canal in detail, it is necessary to say a word about the general features of its composition in the fully-developed man. The mature alimentary canal in man is constructed in all its main features like that of all the higher mammals, and particularly resembles that of the Catarrhines, the narrow-nosed apes of the Old World. The entrance into it, the mouth, is armed with thirty-two teeth, fixed in rows in the upper and lower jaws. As we have seen, our dentition is exactly the same as that of the Catarrhines, and differs from that of all other animals p. 257. Above the mouth-cavity is the double nasal cavity; they are separated by the palate-wall. But we saw that this separation is not there from the first, and that originally there is a common mouth-nasal cavity in the embryo; and this is only divided afterwards by the hard palate into two—the nasal cavity above and that of the mouth below (Fig. 311).
At the back the cavity of the mouth is half closed by the vertical curtain that we call the soft palate, in the middle of which is the uvula. A glance into a mirror with the mouth wide open will show its shape. The uvula is interesting because, besides man, it is only found in the ape. At each side of the soft palate are the tonsils. Through the curved opening that we find
[ 309 ]
underneath the soft palate we penetrate into the gullet or pharynx behind the mouth-cavity. Into this opens on either side a narrow canal (the Eustachian tube), through which there is direct communication with the tympanic cavity of the ear (Fig. 320 e). The pharynx is continued in a long, narrow tube, the œsophagus (sr). By this the food passes into the stomach when masticated and swallowed. Into the gullet also opens, right above, the trachea (lr), that leads to the lungs. The entrance to it is covered by the epiglottis, over which the food slides. The cartilaginous epiglottis is found only in the mammals, and has developed from the fourth branchial arch of the fishes and amphibia. The lungs are found, in man and all the mammals, to the right and left in the pectoral cavity, with the heart between them. At the upper end of the trachea there is, under the epiglottis, a specially differentiated part, strengthened by a cartilaginous skeleton, the larynx. This important organ of human speech also develops from a part of the alimentary canal. In front of the larynx is the thyroid gland, which sometimes enlarges and forms goitre.
The œsophagus descends into the pectoral cavity along the vertebral column, behind the lungs and the heart, pierces the diaphragm, and enters the visceral cavity. The diaphragm is a membrano-muscular partition that completely separates the thoracic from the abdominal cavity in all the mammals (and these alone). This separation is not found in the beginning; there is at first a common breast-belly cavity, the cœloma or pleuro-peritoneal cavity. The diaphragm is formed later on as a muscular horizontal partition between the thoracic and abdominal cavities. It then completely separates the two cavities, and is only pierced by several organs that pass from the one to the other. One of the chief of these organs is the œsophagus. After this has passed through the diaphragm, it expands into the gastric sac in which digestion chiefly takes place. The stomach of the adult man (Fig. 349) is a long, somewhat oblique sac, expanding on the left into a blind sac, the fundus of the stomach (b′), but narrowing on the right, and passing at the pylorus (e) into the small intestine. At this point there is a valve, the pyloric valve (d), between the two sections of the canal; it opens only when the pulpy food passes from the stomach into the intestine. In man and the higher Vertebrates the stomach itself is the chief organ of digestion, and is especially occupied with the solution of the food; this is not the case in many of the lower Vertebrates, which have no stomach, and discharge its function by a part of the gut farther on. The muscular wall of the stomach is comparatively thick; it has externally strong muscles that accomplish the digestive movements, and internally a large quantity of small glands, the peptic glands, which secrete the gastric juice.
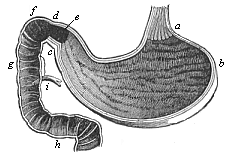 Fig. 349—Human stomach and duodenum, longitudinal section. a cardiac (end of œsophagus), b fundus (blind sac of the left side), c pylorus-fold, d pylorus-valves, e pylorus-cavity, fgh duodenum, i entrance of the gall-duct and the pancreatic duct. (From Meyer.)
Fig. 349—Human stomach and duodenum, longitudinal section. a cardiac (end of œsophagus), b fundus (blind sac of the left side), c pylorus-fold, d pylorus-valves, e pylorus-cavity, fgh duodenum, i entrance of the gall-duct and the pancreatic duct. (From Meyer.) |
Next to the stomach comes the longest section of the alimentary canal, the middle gut or small intestine. Its chief function is to absorb the peptonised fluid mass of food, or the chyle, and it is subdivided into several sections, of which the first (next to the stomach) is called the duodenum (Fig. 349 fgh). It is a short, horseshoe-shaped loop of the gut. The largest glands of the alimentary canal open into it—the liver, the chief digestive gland, that secretes the gall, and the pancreas, which secretes the pancreatic juice. The two glands pour their secretions, the bile and pancreatic juice, close together into the duodenum (i). The opening of the gall-duct is of particular phylogenetic importance, as it is the same in all the Vertebrates, and indicates the principal point of the hepatic or trunk-gut (Gegenbaur). The liver, phylogenetically older than the stomach, is a large gland, rich in blood, in the adult man, immediately under the diaphragm on the left
[ 310 ]
side, and separated by it from the lungs. The pancreas lies a little further back and more to the left. The remaining part of the small intestine is so long that it has to coil itself in many folds in order to find room in the narrow space of the abdominal cavity. It is divided into the jejunum above and the ileum below. In the last section of it is the part of the small intestine at which in the embryo the yelk-sac opens into the gut. This long and thin intestine then passes into the large intestine, from which it is cut off by a special valve. Immediately behind this “Bauhin-valve” the first part of the large intestine forms a wide, pouch-like structure, the cæcum. The atrophied end of the cæcum is the famous rudimentary organ, the vermiform appendix. The large intestine (colon) consists of three parts—an ascending part on the right, a transverse middle part, and a descending part on the left. The latter finally passes through an S-shaped bend into the last section of the alimentary canal, the rectum, which opens behind by the anus. Both the large and small intestines are equipped with numbers of small glands, which secrete mucous and other fluids.
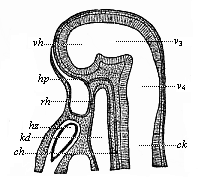 Fig. 350—Median section of the head of a hare-embryo, one-fourth of an inch in length. (From Mihalcovics.) The deep mouth-cleft (hp) is separated by the membrane of the throat (rh) from the blind cavity of the head-gut (kd). hz heart, ch chorda, hp the point at which the hypophysis develops from the mouth-cleft, vh ventricle of the cerebrum, v3, third ventricle (intermediate brain), v4 fourth ventricle (hind brain), ck spinal canal.
Fig. 350—Median section of the head of a hare-embryo, one-fourth of an inch in length. (From Mihalcovics.) The deep mouth-cleft (hp) is separated by the membrane of the throat (rh) from the blind cavity of the head-gut (kd). hz heart, ch chorda, hp the point at which the hypophysis develops from the mouth-cleft, vh ventricle of the cerebrum, v3, third ventricle (intermediate brain), v4 fourth ventricle (hind brain), ck spinal canal. |
For the greater part of its length the alimentary canal is attached to the inner dorsal surface of the abdominal cavity, or to the lower surface of the vertebral column. The fixing is accomplished by means of the thin membranous plate that we call the mesentery.
Although the fully-formed alimentary canal is thus a very elaborate organ, and although in detail it has a quantity of complex structural features into which we cannot enter here, nevertheless the whole complicated structure has been historically evolved from the very simple form of the primitive gut that we find in our gastræad-ancestors, and that every gastrula brings before us to-day. We have already pointed out (Chapter IX) how the epigastrula of the mammals (Fig. 67) can be reduced to the original type of the bell-gastrula, which is now preserved by the amphioxus alone (Fig. 35). Like the latter, the human gastrula and that of all other mammals must be regarded as the ontogenetic reproduction of the phylogenetic form that we call the Gastræa, in which the whole body is nothing but a double-walled gastric sac.
We already know from embryology the manner in which the gut develops in the embryo of man and the other mammals. From the gastrula is first formed the spherical embryonic vesicle filled with fluid (gastrocystis, Fig. 106). In the dorsal wall of this the sole-shaped embryonic shield is developed, and on the under-side of this a shallow groove appears in the middle line, the first trace of the later, secondary alimentary tube. The gut-groove becomes deeper and deeper, and its edges bend towards each other, and finally form a tube.
As we have seen, this simple cylindrical gut-tube is at first completely closed before and behind in man and in the Vertebrates generally (Fig. 148); the permanent openings of the alimentary canal, the mouth and anus, are only formed later on, and from the outer skin. A mouth-pit appears in the skin in front (Fig. 350 hp), and this grows towards the blind fore-end of the cavity of the head-gut (kd), and at length breaks into it. In the same way a shallow anus-pit is formed in the skin behind, which grows deeper and deeper, advances towards the blind hinder end of the pelvic gut, and at last connects with it. There is at first, both before and behind, a thin partition between the external cutaneous pit and the blind end of the gut—the throat-membrane in front and the anus-membrane behind; these disappear when the connection takes place.
Directly in front of the anus-opening the allantois develops from the hind gut; this is the important embryonic structure
[ 311 ]
that forms into the placenta in the Placentals (including man). In this more advanced form the human alimentary canal (and that of all the other mammals) is a slightly bent, cylindrical tube, with an opening at each end, and two appendages growing from its lower wall: the anterior one is the umbilical vesicle or yelk-sac, and the posterior the allantois or urinary sac (Fig. 195).
The thin wall of this simple alimentary tube and its ventral appendages is found, on microscopic examination, to consist of two strata of cells. The inner stratum, lining the entire cavity, consists of larger and darker cells, and is the gut-gland layer. The outer stratum consists of smaller and lighter cells, and is the gut-fibre layer. The only exception is in the cavities of the mouth and anus, because these originate from the skin. The inner coat of the mouth-cavity is not provided by the gut-gland layer, but by the skin-sense layer; and its muscular substratum is provided, not by the gut-fibre, but the skin-fibre, layer. It is the same with the wall of the small anus-cavity.
If it is asked how these constituent layers of the primitive gut-wall are related to the various tissues and organs that we find afterwards in the fully-developed system, the answer is very simple. It can be put in a single sentence. The epithelium of the gut—that is to say, the internal soft stratum of cells that lines the cavity of the alimentary canal and all its appendages, and is immediately occupied with the processes of nutrition—is formed solely from the gut-gland layer; all other tissues and organs that belong to the alimentary canal and its appendages originate from the gut-fibre layer. From the latter is also developed the whole of the outer envelope of the gut and its appendages; the fibrous connective tissue and the smooth muscles that compose its muscular layer, the cartilages that support it (such as the cartilages of the larynx and the trachea), the blood-vessels and lymph-vessels that absorb the nutritive fluid from the intestines—in a word, all that there is in the alimentary system besides the epithelium of the gut. From the same layer we also get the whole of the mesentery, with all the organs embedded in it—the heart, the large blood-vessels of the body, etc.
 Fig. 351—Scales or cutaneous teeth of a shark (Centrophorus calceus). A three-pointed tooth rises obliquely on each of the quadrangular bony plates that lie in the corium. (From Gegenbaur.)
Fig. 351—Scales or cutaneous teeth of a shark (Centrophorus calceus). A three-pointed tooth rises obliquely on each of the quadrangular bony plates that lie in the corium. (From Gegenbaur.) |
Let us now leave this original structure of the mammal gut for a moment, in order to compare it with the alimentary canal of the lower Vertebrates, and of those Invertebrates that we have recognised as man’s ancestors. We find, first of all, in the lowest Metazoa, the Gastræads, that the gut remains permanently in the very simple form in which we find it transitorily in the palingenetic gastrula of the other animals; it is thus in the Gastremaria (Pemmatodiscus), the Physemaria (Prophysema), the simplest Sponges (Olynthus), the freshwater Polyps (Hydra), and the ascula-embryos of many other Cœlenteria (Figs. 233–238). Even in the simplest forms of the Platodes, the Rhabdocœla (Fig. 240), the gut is still a simple straight tube, lined with the entoderm; but with the important difference that in this case its single opening, the primitive mouth (m), has formed a muscular gullet (sd) by invagination of the skin.
We have the same simple form in the gut of the lowest Vermalia (Gastrotricha, Fig. 242, Nematodes, Sagitta, etc.). But in these a second important opening of the gut has been formed at the opposite end to the mouth, the anus (Fig. 242 a).
[ 312 ]
We see a great advance in the structure of the vermalian gut in the remarkable Balanoglossus (Fig. 245), the sole survivor of the Enteropneust class. Here we have the first appearance of the division of the alimentary tube into two sections that characterises the Chordonia. The fore half, the head-gut (cephalogaster), becomes the organ of respiration (branchial gut, Fig. 245 k); the hind half, the trunk-gut (truncogaster), alone acts as digestive organ (hepatic gut, d). The differentiation of these two parts of the gut in the Enteropneust is just the same as in all the Tunicates and Vertebrates.
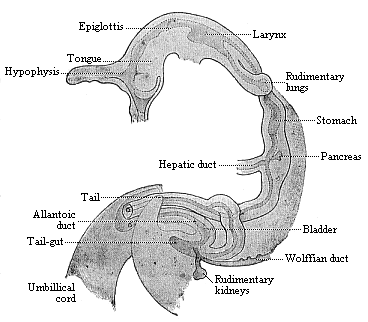 Fig. 352—Gut of a human embryo, one-sixth of an inch long. (From His.)
Fig. 352—Gut of a human embryo, one-sixth of an inch long. (From His.) |
It is particularly interesting and instructive in this connection to compare the Enteropneusts with the Ascidia and the Amphioxus (Figs. 220, 210)—the remarkable animals that form the connecting link between the Invertebrates and the Vertebrates. In both forms the gut is of substantially the same construction; the anterior section forms the respiratory branchial gut, the posterior the digestive hepatic gut. In both it develops palingenetically from the primitive gut of the gastrula, and in both the hinder end of the medullary tube covers the primitive mouth to such an extent that the remarkable medullary intestinal duct is formed, the passing communication between the neural and intestinal tubes (canalis neurentericus, Figs. 83, 85 ne). In the vicinity of the closed primitive mouth, possibly in its place, the later anus is developed. In the same way the mouth is a fresh formation in the Amphioxus and the Ascidia. It is the same with the human mouth and that of the Craniotes generally. The secondary formation of the mouth in the Chordonia is probably connected with the development of the gill-clefts which are formed in the gut-wall immediately behind the mouth. In this way the anterior section of the gut is converted into a respiratory organ. I have already pointed out that this modification is distinctive of the
[ 313 ]
Vertebrates and Tunicates. The phylogenetic appearance of the gill-clefts indicates the commencement of a new epoch in the stem-history of the Vertebrates.
In the further ontogenetic development of the alimentary canal in the human embryo the appearance of the gill-clefts is the most important process. At a very early stage the gullet-wall joins with the external body-wall in the head of the human embryo, and this is followed by the formation of four clefts, which lead directly into the gullet from without, on the right and left sides of the neck, behind the mouth. These are the gill or gullet clefts, and the partitions that separate them are the gill or gullet-arches (Fig. 171). These are most interesting embryonic structures. They show us that all the higher Vertebrates reproduce in their earlier stages, in harmony with the biogenetic law, the process that had so important a part in the rise of the whole Chordonia-stem. This process was the differentiation of the gut into two sections—an anterior respiratory section, the branchial gut, that was restricted to breathing, and a posterior digestive section, the hepatic gut. As we find this highly characteristic differentiation of the gut into two different sections in all the Vertebrates and all the Tunicates, we may conclude that it was also found in their common ancestors, the Prochordonia—especially as even the Enteropneusts have it. (Cf. pp. 119, 151, 227, Figs. 210, 220, 245.) It is entirely wanting in all the other Invertebrates.
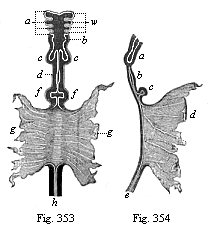 Fig. 353—Gut of a dog-embryo (shown in Fig. 202, from Bischoff), seen from the ventral side. a gill-arches (four pairs), b rudiments of pharynx and larynx, c lungs, d stomach, f liver, g walls of the open yelk-sac (into which the middle gut opens with a wide aperture), h rectum.
Fig. 353—Gut of a dog-embryo (shown in Fig. 202, from Bischoff), seen from the ventral side. a gill-arches (four pairs), b rudiments of pharynx and larynx, c lungs, d stomach, f liver, g walls of the open yelk-sac (into which the middle gut opens with a wide aperture), h rectum.
Fig. 354—The same gut seen from the right. a lungs, b stomach, c liver, d yelk-sac, e rectum.) |
There is at first only one pair of gill-clefts in the Amphioxus, as in the Ascidia and Enteropneusts; and the Copelata (Fig. 225) have only one pair throughout life. But the number presently increases in the former. In the Craniotes, however, it decreases still further. The Cyclostomes have six to eight pairs (Fig. 247); some of the Selachii six or seven pairs, most of the fishes only four or five pairs. In the embryo of man, and the higher Vertebrates generally, where they make an appearance at an early stage, only three or four pairs are developed. In the fishes they remain throughout life, and form an exit for the water taken in at the mouth (Figs. 249–251). But they are partly lost in the amphibia, and entirely in the higher Vertebrates. In these nothing is left but a relic of the first gill-cleft. This is formed into a part of the organ of hearing; from it are developed the external meatus, the tympanic cavity, and the Eustachian tube. We have already considered these remarkable structures, and need only point here to the interesting fact that our middle and external ear is a modified inheritance from the fishes. The branchial arches also, which separate the clefts, develop into very different parts. In the fishes they remain gill-arches, supporting the respiratory gill-leaves. It is the same with the lowest amphibia, but in the higher amphibia they undergo various modifications; and in the three higher classes of Vertebrates (including man) the hyoid bone and the ossicles of the ear develop from them. (Cf. p. 291.)
From the first gill-arch, from the inner surface of which the muscular tongue proceeds, we get the first structure of the maxillary skeleton—the upper and lower jaws, which surround the mouth and support the teeth. These important parts are wholly wanting in the two lowest classes of Vertebrates, the Acrania and Cyclostoma. They appear first in the earliest Selachii (Figs. 248–251), and have been transmitted from this stem-group of the Gnathostomes to the higher
[ 314 ]
Vertebrates. Hence the original formation of the skeleton of the mouth can be traced to these primitive fishes, from which we have inherited it. The teeth are developed from the skin that clothes the jaws. As the whole mouth cavity originates from the outer integument (Fig. 350), the teeth also must come from it. As a fact, this is found to be the case on microscopic examination of the development and finer structure of the teeth. The scales of the fishes, especially of the shark type (Fig. 351), are in the same position as their teeth in this respect (Fig. 252). The osseous matter of the tooth (dentine) develops from the corium; its enamel covering is a secretion of the epidermis that covers the corium. It is the same with the cutaneous teeth or placoid scales of the Selachii. At first the whole of the mouth was armed with these cutaneous teeth in the Selachii and in the earliest amphibia. Afterwards the formation of them was restricted to the edges of the jaws.
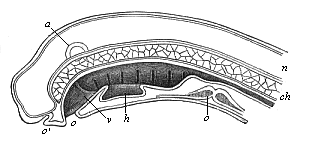 |
Fig. 355—Median section of the head of a Petromyzon-larva. (From Gegenbaur.) h hypobranchial groove (above it in the gullet we see the internal openings of the seven gill-clefts), v velum, o mouth, c heart, a auditory vesicle, n neural tube, ch chorda. |
Hence our human teeth are, in relation to their original source, modified fish-scales. For the same reason we must regard the salivary glands, which open into the mouth, as epidermic glands, as they are formed, not from the glandular layer of the gut like the rest of the alimentary glands, but from the epidermis, from the horny plate of the outer germinal layer. Naturally, in harmony with this evolution of the mouth, the salivary glands belong genetically to one series with the sudoriferous, sebaceous, and mammary glands.
Thus the human alimentary canal is as simple as the primitive gut of the gastrula in its original structure. Later it resembles the gut of the earliest Vermalia (Gastrotricha). It then divides into two sections, a fore or branchial gut and a hind or hepatic gut, like the alimentary canal of the Balanoglossus, the Ascidia, and the Amphioxus. The formation of the jaws and the branchial arches changes it into a real fish-gut (Selachii). But the branchial gut, the one reminiscence of our fish-ancestors, is afterwards atrophied as such. The parts of it that remain are converted into entirely different structures.
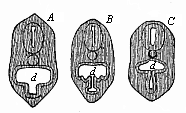 Fig. 356—Transverse section of the head of a Petromyzon-larva. (From Gegenbaur.) Beneath the pharynx (d) we see the hypobranchial groove; above it the chorda and neural tube. A, B, C stages of constriction.
Fig. 356—Transverse section of the head of a Petromyzon-larva. (From Gegenbaur.) Beneath the pharynx (d) we see the hypobranchial groove; above it the chorda and neural tube. A, B, C stages of constriction. |
But, although the anterior section of our alimentary canal thus entirely loses its original character of branchial gut, it retains the physiological character of respiratory gut. We are now astonished to find that the permanent respiratory organ of the higher Vertebrates, the air-breathing lung, is developed from this first part of the alimentary canal. Our lungs, trachea, and larynx are formed from the ventral wall of the branchial gut. The whole of the respiratory apparatus, which occupies the greater part of the pectoral cavity in the adult man, is at first merely a small pair of vesicles or sacs, which grow out of the floor of the head-gut immediately behind the gills (Figs. 354 c, 147 l). These vesicles are found in all the Vertebrates except the two lowest classes, the Acrania and Cyclostomes. In the lower Vertebrates they do not develop
[ 315 ]
into lungs, but into a large air-filled bladder, which occupies a good deal of the body-cavity and has a quite different purport. It serves, not for breathing, but to effect swimming movements up and down, and so is a sort of hydrostatic apparatus—the floating bladder of the fishes (nectocystis, p. 233). However, the human lungs, and those of all air-breathing Vertebrates, develop from the same simple vesicular appendage of the head-gut that becomes the floating bladder in the fishes.
At first this bladder has no respiratory function, but merely acts as hydrostatic apparatus for the purpose of increasing or lessening the specific gravity of the body. The fishes, which have a fully-developed floating bladder, can press it together, and thus condense the air it contains. The air also escapes sometimes from the alimentary canal, through an air-duct that connects the floating bladder with the pharynx, and is ejected by the mouth. This lessens the size of the bladder, and so the fish becomes heavier and sinks. When it wishes to rise again, the bladder is expanded by relaxing the pressure. In many of the Crossopterygii the wall of the bladder is covered with bony plates, as in the Triassic Undina (Fig. 254).
This hydrostatic apparatus begins in the Dipneusts to change into a respiratory organ; the blood-vessels in the wall of the bladder now no longer merely secrete air themselves, but also take in fresh air through the air-duct. This process reaches its full development in the Amphibia. In these the floating bladder has turned into lungs, and the air-passage into a trachea. The lungs of the Amphibia have been transmitted to the three higher classes of Vertebrates. In the lowest Amphibia the lungs on either side are still very simple transparent sacs with thin walls, as in the common water-salamander, the Triton. It still entirely resembles the floating bladder of the fishes. It is true that the Amphibia have two lungs, right and left. But the floating bladder is also double in many of the fishes (such as the early Ganoids), and divides into right and left halves. On the other hand, the lung is single in Ceratodus (Fig. 257).
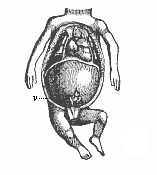 Fig. 357—Thoracic and abdominal viscera of a human embryo of twelve weeks. (From Kölliker.) The head is omitted. Ventral and pectoral walls are removed. The greater part of the body-cavity is taken up with the liver, from the middle part of which the cæcum and the vermiform appendix protrude. Above the diaphragm, in the middle, is the conical heart; to the right and left of it are the two small lungs.
Fig. 357—Thoracic and abdominal viscera of a human embryo of twelve weeks. (From Kölliker.) The head is omitted. Ventral and pectoral walls are removed. The greater part of the body-cavity is taken up with the liver, from the middle part of which the cæcum and the vermiform appendix protrude. Above the diaphragm, in the middle, is the conical heart; to the right and left of it are the two small lungs. |
In the human embryo and that of all the other Amniotes the lungs develop from the hind part of the ventral wall of the head-gut (Fig. 149). Immediately behind the single structure of the thyroid gland a median groove, the rudiment of the trachea, is detached from the gullet. From its hinder end a couple of vesicles develop—the simple tubular rudiments of the right and left lungs. They afterwards increase considerably in size, fill the greater part of the thoracic cavity, and take the heart between them. Even in the frogs we find that the simple sac has developed into a spongy body of peculiar froth-like tissue. The originally short connection of the pulmonary sacs with the head-gut extends into a long, thin tube. This is the wind-pipe (trachea); it opens into the gullet above, and divides below into two branches which go to the two lungs. In the wall of the trachea circular cartilages develop, and these keep it open. At its upper end, underneath its pharyngeal opening, the larynx is formed—the organ of voice and speech. The larynx is found at various stages of development in the Amphibia, and comparative anatomists are in a position to trace the progressive growth of this important organ from the rudimentary structure of the lower Amphibia up to the elaborate and delicate vocal apparatus that we have in the larynx of man and of the birds.
We must refer here to an interesting rudimentary organ of the respiratory gut, the thyroid gland, the large gland in front of the larynx, that lies below the “Adam’s
[ 316 ]
apple,” and is often especially developed in the male sex. It has a certain function—not yet fully understood—in the nutrition of the body, and arises in the embryo by constriction from the lower wall of the pharynx. In many mining districts the thyroid gland is peculiarly liable to morbid enlargement, and then forms goitre, a growth that hangs at the front of the neck. But it is much more interesting phylogenetically. As Wilhelm Müller, of Jena, has shown, this rudimentary organ is the last relic of the hypobranchial groove, which we considered in a previous chapter, and which runs in the middle line of the gill-crate in the Ascidia and Amphioxus, and conveys food to the stomach. (Cf. p. 184, Fig. 246). We still find it in its original character in the larvæ of the Cyclostomes (Figs. 355, 356).
The second section of the alimentary canal, the trunk or hepatic gut, undergoes not less important modifications among our vertebrate ancestors than the first section. In tracing the further development of this digestive part of the gut, we find that most complex and elaborate organs originate from a very rudimentary original structure. For clearness we may divide the digestive gut into three sections: the fore gut (with œsophagus and stomach), the middle gut (duodenum, with liver, pancreas, jejunum, and ileum, and the hind gut (colon and rectum). Here again we find vesicular growths or appendages of the originally simple gut developing into a variety of organs. Two of these embryonic structures, the yelk-sac and allantois, are already known to us. The two large glands that open into the duodenum, the liver and pancreas, are growths from the middle and most important part of the trunk-gut.
Immediately behind the vesicular rudiments of the lungs comes the section of the alimentary canal that forms the stomach (Figs. 353 d, 354 b). This sac-shaped organ, which is chiefly responsible for the solution and digestion of the food, has not in the lower Vertebrates the great physiological importance and the complex character that it has in the higher. In the Acrania and Cyclostomes and the earlier fishes we can scarcely distinguish a real stomach; it is represented merely by the short piece from the branchial to the hepatic gut. In some of the other fishes also the stomach is only a very simple spindle-shaped enlargement at the beginning of the digestive section of the gut, running straight from front to back in the median plane of the body, underneath the vertebral column. In the mammals its first structure is just as rudimentary as it is permanently in the preceding. But its various parts soon begin to develop. As the left side of the spindle-shaped sac grows much more quickly than the right, and as it turns considerably on its axis at the same time, it soon comes to lie obliquely. The upper end is more to the left, and the lower end more to the right. The foremost end draws up into the longer and narrower canal of the œsophagus. Underneath this on the left the blind sac (fundus) of the stomach bulges out, and thus the later form gradually develops (Figs. 349, 184 e). The original longitudinal axis becomes oblique, sinking below to the left and rising to the right, and approaches nearer and nearer to a transverse position. In the outer layer of the stomach-wall the powerful muscles that accomplish the digestive movements develop from the gut-fibre layer. In the inner layer a number of small glandular tubes are formed from the gut-gland layer; these are the peptic glands that secrete the gastric juice. At the lower end of the gastric sac is developed the valve that separates it from the duodenum (the pylorus, Fig. 349 d).
Underneath the stomach there now develops the disproportionately long stretch of the small intestine. The development of this section is very simple, and consists essentially in an extremely rapid and considerable growth lengthways. It is at first very short, quite straight, and simple. But immediately behind the stomach we find at an early stage a horseshoe-shaped bend and loop of the gut, in connection with the severance of the alimentary canal from the yelk-sac and the development of the first mesentery. The thin delicate membrane that fastens this loop to the ventral side of the vertebral column, and fills the inner bend of the horseshoe formation, is the first rudiment of the mesentery (Fig. 147 g). We find at an early stage a considerable growth of the small intestine; it is thus forced to coil itself in a number of loops. The various sections that we have to distinguish in it are differentiated in a very simple way—the duodenum (next to the stomach), the succeeding long jejunum, and the last section of the small intestine, the ileum.
[ 317 ]
From the duodenum are developed the two large glands that we have already mentioned—the liver and pancreas. The liver appears first in the shape of two small sacs, that are found to the right and left immediately behind the stomach (Figs. 353 f, 354 c). In many of the lower Vertebrates they remain separate for a long time (in the Myxinoides throughout life), or are only imperfectly joined. In the higher Vertebrates they soon blend more or less completely to form a single large organ. The growth of the liver is very brisk at first. In the human embryo it grows so much in the second month of development that in the third it occupies by far the greater part of the body-cavity (Fig. 357). At first the two halves develop equally; afterwards the left falls far behind the right. In consequence of the unsymmetrical development and turning of the stomach and other abdominal viscera, the whole liver is now pushed to the right side. Although the liver does not afterwards grow so disproportionately, it is comparatively larger in the embryo at the end of pregnancy than in the adult. Its weight relatively to that of the whole body is 1 : 36 in the adult, and 1 : 18 in the embryo. Hence it is very important physiologically during embryonic life; it is chiefly concerned in the formation of blood, not so much in the secretion of bile.
Immediately behind the liver a second large visceral gland develops from the duodenum, the pancreas or sweetbread. It is wanting in most of the lowest classes of Vertebrates, and is first found in the fishes. This organ is also an outgrowth from the gut.
The last section of the alimentary canal, the large intestine, is at first in the embryo a very simple, short, and straight tube, which opens behind by the anus. It remains thus throughout life in the lower Vertebrates. But it grows considerably in the mammals, coils into various folds, and divides into two sections, the first and longer of which is the colon, and the second the rectum. At the beginning of the colon there is a valve (valvula Bauhini) that separates it from the small intestine. Immediately behind this there is a sac-like growth, which enlarges into the cæcum (Fig. 357 v). In the plant-eating mammals this is very large, but it is very small or completely atrophied in the flesh-eaters. In man, and most of the apes, only the first portion of the cæcum is wide; the blind end-part of it is very narrow, and seems later to be merely a useless appendage of the former. This “vermiform appendage” is very interesting as a rudimentary organ. The only significance of it in man is that not infrequently a cherry-stone or some other hard and indigestible matter penetrates into its narrow cavity, and by setting up inflammation and suppuration causes the death of otherwise sound men. Teleology has great difficulty in giving a rational explanation of, and attributing to a beneficent Providence, this dreaded appendicitis. In our plant-eating ancestors this rudimentary organ was much larger and had a useful function.
Finally, we have important appendages of the alimentary tube in the bladder and urethra, which belong to the alimentary system. These urinary organs, acting as reservoir and duct for the urine excreted by the kidneys, originate from the innermost part of the allantoic pedicle. In the Dipneusts and Amphibia, in which the allantoic sac first makes its appearance, it remains within the body-cavity, and functions entirely as bladder. But in all the Amniotes it grows far outside of the body-cavity of the embryo, and forms the large embryonic “primitive bladder,” from which the placenta develops in the higher mammals. This is lost at birth. But the long stalk or pedicle of the allantois remains, and forms with its upper part the middle vesico-umbilical ligament, a rudimentary organ that goes in the shape of a solid string from the vertex of the bladder to the navel. The lowest part of the allantoic pedicle (or the “urachus”) remains hollow, and forms the bladder. At first this opens into the last section of the gut in man as in the lower Vertebrates; thus there is a real cloaca, which takes off both urine and excrements. But among the mammals this cloaca is only permanent in the Monotremes, as it is in all the birds, reptiles, and amphibia. In all the other mammals (marsupials and placentals) a transverse partition is afterwards formed, and this separates the urogenital aperture in front from the anus-opening behind. (Cf. p. 249 and Chapter 29.)
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XXVI
Chapter XXVIII
Figs. 1–209
Figs. 210–408