Scarcely less striking than the example of the air-breathing Crabs, is the behaviour of the heart in the great section Edriophthalma, which may advantageously be divided, after the example of Dana and Spence Bate, only into two orders, the Amphipoda and the Isopoda.
In the Amphipoda, to which the above-mentioned naturalists correctly refer the Caprellidæ and Cyamidæ (Latreille’s Læmodipoda), the heart has always the same position; it extends in the form of a long tube through the six segments following the head, and has three pairs of fissures, furnished with valves, for the entrance of the blood, situated in the second, third, and fourth of these segments. It was found to be of this structure by La Valette in Niphargus (Gammarus puteanus), and by Claus in Phronima; and I have found it to be the same in a considerable number of species belonging to the most different families.1
The sole unimportant exception which I have hitherto met with is presented by the genus Brachyscelus,2 in which the heart possesses only two pairs of fissures, as it extends forward only into the second body-segment, and is destitute of the pair of fissures situated in this segment in other forms.3
Considering this uniformity presented by the heart in the entire order of the Amphipoda, it cannot but seem very remarkable, that in the very next order of the Isopoda, we find it to be one of the most changeable organs.
In the cheliferous Isopods (Tanais) the heart resembles that of the Amphipoda in its elongated tubular form, as well as in the number and position of the fissures, but with this difference, that the two fissures of each pair do not lie directly opposite each other.
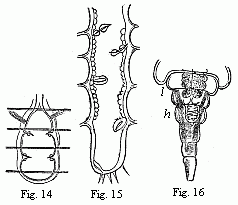
Fig. 14. Heart of a young Cassidina. Fig. 15. Heart of a young Anilocra.
Fig. 16. Abdomen of the male of Entoniscus Cancrorum. h. Heart. l. Liver.
In all other Isopoda the heart is removed towards the abdomen. In the wonderfully deformed parasitic Isopods of the Porcellanæ (Entoniscus porcellanæ), the spherical heart of the female is confined to a short space of the elongated first abdominal segment, and seems to possess only a single pair of fissures. In the male of Entoniscus Cancrorum (n. sp.), the heart (Fig. 16) is situated in the third abdominal segment. In the Cassidinæ, the heart (Fig. 14) is likewise short and furnished with two pairs of fissures, situated in the last segment of the thorax and the first segment of the abdomen. Lastly, in a young Anilocra, I find the heart (Fig. 15) extending through the whole length of the abdomen and furnished with four (or five?) fissures, which are not placed in pairs but alternately to the right and left in successive segments. In other animals of this order, which I have as yet only cursorily examined, further differences will no doubt occur. But why, in two orders so nearly allied to each other, should we find in the one such a constancy, in the other such a variability, of the same highly important organ? From the schoolmen we need expect no explanation, they will either decline the discussion of the “wherefore” as foreign to their province, as lying beyond the boundaries of Natural History, or seek to put down the importunate question by means of a sounding paraphrase of the facts, abundantly sprinkled with Greek words. As I have unfortunately forgotten my Greek, the second way out of the difficulty is closed to me; but as I luckily reckon myself not amongst the incorporated masters, but, to use Baron von Liebig’s expression, amongst the “promenaders on the outskirts of Natural History,” this affected hesitation of the schoolmen cannot dissuade me from seeking an answer, which indeed presents itself most naturally from Darwin’s point of view.
As not only the Tanaides (which reasons elsewhere stated (vide suprà) justify us in regarding as particularly nearly related to the primitive Isopod) and the Amphipoda, but also the Decapod Crustacea, possess a heart with three pairs of fissures essentially in the same position; and as the same position of the heart recurs (vide infrà) even in the embryos of the Mantis-Shrimps (Squilla), in which the heart of the adult animal, and even, as I have elsewhere shown, that of the larvæ when still far from maturity, extends in the form of a long tube with numerous openings far into the abdomen, we must unhesitatingly regard the heart of the Amphipoda as the primitive form of that organ in the Edriophthalma. As, moreover, in these animals the blood flows from the respiratory organs to the heart without vessels, it is very easy to see how advantageous it must be to them to have these organs as much approximated as possible. We have reason to regard as the primitive mode of respiration, that occurring in Tanais (vide suprà). Now, where, as in the majority of the Isopoda, branchiæ were developed upon the abdomen, the position and structure of the heart underwent a change, as it approached them more nearly, but without the reproduction of a common plan for these earlier modes of structure, either because this transformation of the heart took place only after the division of the primary form into subordinate groups, or because, at least at the time of this division, the varying heart had not yet become fixed in any new form. Where, on the contrary, respiration remained with the anterior part of the body,—whether in the primitive fashion of Zoëa, as in the Tanaides, or by the development of branchiæ on the thorax, as in the Amphipoda,—the primitive form of the heart was inherited unchanged, because any variations which might make their appearance were rather injurious than advantageous, and disappeared again immediately.
I close this series of isolated examples with an observation which indeed only half belongs to the province of the Crustacea to which these pages ought to be confined, and which also has no further connexion with the preceding circumstances than that of being an “intelligible and intelligence-bringing fact” only from the point of view of Darwin’s theory. To-day as I was opening a specimen of Lepas anatifera in order to compare the animal with the description in Darwin’s ‘Monograph on the Subclass Cirripedia,’ I found in the shell of this Cirripede, a blood-red Annelide, with a short, flat body, about half an inch long and two lines in breadth, with twenty-five body-segments, and without projecting setigerous tubercles or jointed cirri. The small cephalic lobe bore four eyes and five tentacles; each body-segment had on each side at the margin a tuft of simple setæ directed obliquely upwards, and at some distance from this, upon the ventral surface, a group of thicker setæ with a strongly uncinate bidentate apex. There was above each of the lateral tufts of bristles a branchia, simple on a few of the foremost segments, and then strongly arborescent to the end of the body. The animal, a female filled with ova, evidently, from these characters, belongs to the family of the Amphinomidæ; the only family the members of which, being excellent swimmers, live in the open sea.
That this animal had not strayed accidentally into the Lepas, but appertained to it as a regular and permanent guest, is evidenced by its considerable size in proportion to the narrow entrance of the test of the Lepas, by the complete absence of the iridescence which usually distinguishes the skin of free Annelides and especially of the Amphinomidæ, by the formation and position of the inferior setæ, etc. But that a worm belonging to this particular family Amphinomidæ living in the high sea, occurs as a guest in the Lepas, which also floats in the sea attached to wood, etc., is at once intelligible from the stand-point of the Darwinian theory, whilst the relationship of this parasite to the free-living worms of the open sea remains perfectly unintelligible under the supposition that it was independently created for dwelling in the Lepas.
But however favourable the examples hitherto referred to may be for Darwin, the objection may be raised against them, and that with perfect justice, that they are only isolated facts, which, when the considerations founded upon them are carried far beyond what is immediately given, may only too easily lead us from the right path, with the deceptive glimmer of an ignis fatuus. The higher the structure to be raised, the wider must be the assuring base of well-sifted facts.
Let us turn then to a wider field, that of the developmental history of the Crustacea, upon which science has already brought together a varied abundance of remarkable facts, which, however, have remained a barren accumulation of unmanageable raw-material, and let us see how, under Darwin’s hand, these scattered stones unite to form a well-jointed structure, in which everything, bearing and being borne, finds its significant place. Under Darwin’s hand! for I shall have nothing to do except just to place the building stones in the position which his theory indicates for them. “When kings build, the carters have to work.”
1. The young animals in the egg, a little before their exclusion, are usually particularly convenient for the observation of the fissures in the heart; they are generally sufficiently transparent, the movements of the heart are less violent than at a later period, and they lie still even without the pressure of a glass cover. Considering the common opinion as to the distribution of the Amphipoda, namely, that they increase in multiplicity towards the poles, and diminish towards the equator, it may seem strange that I speak of a considerable number of species on a subtropical coast. I therefore remark that in a few months and without examining any depths inaccessible from the shore, I obtained 38 different species, of which 34 are new, which, with the previously known species (principally described by Dana) gives 60 Brazilian Amphipoda, whilst Kröyer in his ‘Grönlands Amfipoder’ was acquainted with only 28 species, including 2 Læmodipoda, from the Arctic Seas, although these had been investigated by a far greater number of Naturalists.
2. According to Milne-Edwards’ arrangement the females of this genus would belong to the “Hypérines ordinaires” and the previously unknown males to the “Hypérines anormales,” the distinguishing character of which, namely the curiously zigzagged inferior antennæ, is only a sexual peculiarity of the male animals. In systematising from single dead specimens, as to the sex, age, etc. of which nothing is known, similar errors are unavoidable. Thus, in order to give another example of very recent date, a celebrated Ichthyologist, Bleeker, has lately distinguished two groups of the Cyprinodontes as follows: some, the Cyprinodontini, have a “pinna analis non elongata,” and the others, the Aplocheilini, a “pinna analis elongata”: according to this the female of a little fish which is very abundant here would belong to the first, and the male to the second group. Such mistakes, as already stated, are unavoidable by the “dry-skin” philosopher, and therefore excusable; but they nevertheless prove in how random a fashion the present systematic zoology frequently goes on, without principles or sure foundations, and how much it is in want of the infallible touchstone for the value of the different characters, which Darwin’s theory promises to furnish.
3. I find, in Milne-Edwards’ ‘Leçons sur la Physiol. et l’Anat. comp.’ 3 page 197, the statement that, according to Frey and Leuckart, the heart of Caprella linearis possesses five pairs of fissures. I have examined perfectly transparent young Caprellæ (probably the young of Caprella attenuata, Dana, with which they occurred), but can only find the usual three pairs.