Chapter III
MORPHOLOGY OF CRUSTACEA—NAUPLIUS-LARVÆ
If the absence of contradictions among the inferences deduced from them for a narrow and consequently easily surveyed department must prepossess us in favour of Darwin’s views, it must be welcomed as a positive triumph of his theory if far-reaching conclusions founded upon it should subsequently be confirmed by facts, the existence of which science, in its previous state, by no means allowed us to suspect. From many results of this kind upon which I could report, I select as examples, two, which were of particular importance to me, and relate to discoveries the great significance of which in the morphology and classification of the Crustacea will not be denied even by the opponents of Darwin.
Considerations upon the developmental history of the Crustacea had led me to the conclusion that, if the higher and lower Crustacea were at all derivable from common progenitors, the former also must once have passed through Nauplius-like conditions. Soon afterwards I discovered Naupliiform larvæ of Shrimps (‘Archiv für Naturgeschichte’ 1860, i, p. 8), and I must admit that this discovery gave me the first decided turn in Darwin’s favour.
The similar number of segments1 occurring in the Crabs and Macrura, Amphipoda and Isopoda, in which the last seven segments are always different from the preceding ones in the appendages with which they are furnished, could only be regarded as an inheritance from the same ancestors. And if at the present day the majority of the Crabs and Macrura, and indeed the Stalk-eyed Crustacea in general, pass through Zoëa-like developmental states, and the same mode of transformation was to be ascribed to their ancestors, the same thing must also apply, if not to the immediate ancestors of the Amphipoda and Isopoda, at least to the common progenitors of these and the Stalk-eyed Crustacea. Any such assumption as this was, however, very hazardous, so long as not a single fact properly relating to the Edriophthalma could be adduced in its support, as the structure of this very coherent group seemed to be almost irreconcilable with many peculiarities of the Zoëa. Thus, in my eyes, this point long constituted one of the chief difficulties in the application of the Darwinian views to the Crustacea, and I could scarcely venture to hope that I might yet find traces of this passage through the Zoëa-form among the Amphipoda or Isopoda, and thus obtain a positive proof of the correctness of this conclusion. At this point Van Beneden’s statement that a cheliferous Isopod (Tanais Dulongii), belonging, according to Milne-Edwards, to the same family as the common Asellus aquaticus, possesses a carapace like the Decapoda, directed my attention to these animals, and a careful examination proved that these Isopods have preserved, more truly than any other adult Crustacea, many of the most essential peculiarities of the Zoëae, especially their mode of respiration. Whilst in all other Oniscoida the abdominal feet serve for respiration, these in our cheliferous Isopod (Fig. 2) are solely motory organs, into which no blood-corpuscle ever enters, and the chief seat of respiration is, as in the Zoëae, in the lateral parts of the carapace, which are abundantly traversed by currents of blood, and beneath which a constant stream of water passes, maintained, as in Zoëae and the adult Decapoda, by an appendage of the second pair of maxillæ, which is wanting in all other Edriophthalma.
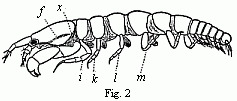
Fig. 2. Tanais dubius (?) Kr. hermaphrodite, magnified, showing the orifice of entrance (x) into the cavity overarched by the carapace, in which an appendage of the second pair of maxillæ (f) plays. On four feet (i, k, l, m) are the rudiments of the lamellæ which subsequently form the brood-cavity.
For both these discoveries, it may be remarked in passing, science is indebted less to a happy chance than immediately to Darwin’s theory.
Species of Penëus live in the European seas, as well as here, and their Nauplius-brood has no doubt repeatedly passed unnoticed through the hands of the numerous naturalists who have investigated those seas, as well as through my own,2 for it has nothing which could attract particular attention amongst the multifarious and often wonderful Nauplius-forms. When I, fancying from the similarity of its movements that it was a young Penëus-Zoëa, had for the first time captured such a larva, and on bringing it under the microscope found a Nauplius differing toto cœlo from this Zoëa, I might have thrown it aside as being completely foreign to the developmental series which I was tracing, if the idea of early Naupliiform stages of the higher Crustacea, which indeed I did not believe to be still extant, had not at the moment vividly occupied my attention.
And if I had not long been seeking among the Edriophthalma for traces of the supposititious Zoëa-state, and seized with avidity upon everything that promised to made this refractory Order serviceable to me, Van Beneden’s short statement could hardly have affected me so much in the manner of an electric shock, and impelled me to a renewed study of the Tanaides, especially as I had once before plagued myself with them in the Baltic, without getting any further than my predecessors, and I have not much taste for going twice over the same ground.
1. Like Claus I do not regard the eyes of the Crustacea as limbs, and therefore admit no ocular segment; on the other hand I count in the median piece of the tail, to which the character of a segment is often denied. In opposition to its interpretation as a segment of the body, only the want of limbs can be cited; in its favour we have the relation of the intestine, which usually opens in this piece, and sometimes even traverses its whole length, as in Microdeutopus and some other Amphipoda. In Microdeutopus, as Spence Bate has already pointed out, one is even led to regard small processes of this tubular caudal piece as rudimentary members. Bell also (‘British Stalk-eyed Crustacea’ p. xx), states that he observed limbs of the last segment in Palæmon serratus in the form of small moveable points.
The attempt has often been made to divide the body of the higher Crustacea into small sections composed of equal numbers of segments, these sections consisting of 3, 5 or 7 segments. None of these attempts has ever met with general acceptance; my own investigations lead me to a conception which nearly approaches Van Beneden’s. I assume four sections of 5 segments each—the primitive body, the fore-body, the hind-body, and the middle-body. The primitive body includes the segments which the naupliiform larva brings with it out of the egg; it is afterwards divided, by the younger sections which become developed in its middle, into the head and tail. To this primitive body belong the two pairs of antennæ, the mandibles and the caudal feet (“posterior pair of pleopoda,” Sp. B.). Even in the mature animal the fact that these terminal sections belong to one another is sometimes betrayed by the resemblance of their appendages, especially that of the outer branch of the caudal feet, with the outer branch (the so-called scale) of the second pair of antennæ. Like the antennæ, the caudal feet may also become the bearers of high sensorial apparatus, as is shown by the ear of Mysis.
The sequence of the sections of the body in order of time seems originally to have been, that first the fore-body, then the hind-body, and finally the middle-body was formed. The fore-body appears, in the adult animal, to be entirely or partially amalgamated with the head; its appendages (siagonopoda Westw.) are all or in part serviceable for the reception of food, and generally sharply distinguished from those of the following group. The segments of the middle-body seem always to put forth limbs immediately after their own appearance, whilst the segments of the hind-body often remain destitute of feet through long portions of the larval life or even throughout life (as in many female Diastylidæ), a reason, among many others, for not, as is usual, regarding the middle-body of the Crustacea as equivalent to the constantly footless abdomen of Insects. The appendages of the middle-body (pereiopoda) seem never, even in their youngest form, to possess two equal branches, a peculiarity which usually characterises the appendages of the hind-body. This is a circumstance which renders very doubtful the equivalence of the middle-body of the Malacostraca with the section of the body which in the Copepoda bears the swimming feet and in the Cirripedia the cirri.
The comprehension of the feet of the hind-body and tail in a single group (as “fausses pattes abdominales,” or as “pleopoda”) seems not to be justifiable. When there is a metamorphosis, they are probably always produced at different periods, and they are almost always quite different in structure and function. Even in the Amphipoda, in which the caudal feet usually resemble in appearance the last two pairs of abdominal feet, they are in general distinguished by some sort of peculiarity, and whilst the abdominal feet are reproduced in wearisome uniformity throughout the entire order, the caudal feet are, as is well-known, amongst the most variable parts of the Amphipoda.
2. Mecznikow has recently found Naupliiform shrimp-larvæ in the sea near Naples.
Chapter II
Chapter IV