The Project Gutenberg EBook of Introduction to the Study of Palæontological Botany, by John Hutton Balfour This eBook is for the use of anyone anywhere in the United States and most other parts of the world at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org. If you are not located in the United States, you'll have to check the laws of the country where you are located before using this ebook. Title: Introduction to the Study of Palæontological Botany Author: John Hutton Balfour Release Date: January 9, 2016 [EBook #50882] Language: English Character set encoding: UTF-8 *** START OF THIS PROJECT GUTENBERG EBOOK INTRODUCTION--PALAEONTOLOGICAL BOTANY *** Produced by Brian Coe, John Campbell and the Online Distributed Proofreading Team at http://www.pgdp.net (This file was produced from images generously made available by The Internet Archive/American Libraries.)
TRANSCRIBER'S NOTE
Basic fractions are displayed as ½ ⅓ ¼ etc; other fractions are shown in the form a/b, eg 3/11 or 13/(34×2).
The caption for an illustration is displayed as a sidenote in the etext. It was shown as a page footnote in the original text.
Obvious typographical errors and punctuation errors have been corrected after careful comparison with other occurrences within the text and consultation of external sources.
More detail can be found at the end of the book.
INTRODUCTION
TO THE STUDY OF
PALÆONTOLOGICAL BOTANY
BY
JOHN HUTTON BALFOUR, A.M. M.D. EDIN.
F.R.S., SEC. R.S.E., F.L.S.
PROFESSOR OF MEDICINE AND BOTANY IN THE UNIVERSITY OF EDINBURGH,
REGIUS KEEPER OF THE ROYAL BOTANIC GARDEN,
AND QUEEN'S BOTANIST FOR SCOTLAND
WITH FOUR LITHOGRAPHIC PLATES, AND UPWARDS OF
ONE HUNDRED WOODCUTS
EDINBURGH
ADAM AND CHARLES BLACK
1872
Printed by R. & R. Clark, Edinburgh.
TO
PROF. HEINRICH ROBERT GOEPPERT, M.D.,
DIRECTOR OF THE BOTANIC GARDEN, BRESLAU;
ONE OF THE MOST EMINENT PALÆONTOLOGICAL
BOTANISTS OF EUROPE,
The following Treatise
IS DEDICATED, WITH BEST RESPECTS, BY HIS
OBLIGED FRIEND
THE AUTHOR.
The subject of Fossil Botany or Palæophytology has formed a part of the Course of Botany in the University of Edinburgh for the last twenty-five years, and the amount of time devoted to the exposition of it has increased. The recent foundation of a Chair of Geology and of a Falconer Palæontological Fellowship in the University seems to require from the Professors of Zoology and Botany special attention to the bearings of their departments of science on the structure of the animals and plants of former epochs of the Earth's history. No one can be competent to give a correct decision in regard to Fossils, unless he has studied thoroughly the present Fauna and Flora of the globe. To give a well-founded opinion in regard to extinct beings, it is essential that the observer should be conversant with the conformation and development of the living ones now on the earth; with their habits, modes of existence and reproduction, the microscopic structure of their tissues, their distribution, and their relation to soil, the atmosphere, temperature, and climate.
There can be no doubt that to become a good Fossil Geologist a student must begin with living animals and plants. The study of Geology must be shared by the[viii] Petralogist, who looks at the condition of the rocks of the globe, the minerals forming them, and their mode of formation; the Chemist, who determines the materials which enter into the composition of minerals and rocks; the Naturalist, who examines the plants and animals found in the various strata; and perhaps also the Natural Philosopher, who calculates from independent sources the phases of the Earth's history. It may be said thus to combine all these students of Science in one brotherhood. Much has been done by the efforts of such men as Hutton and Werner, who were engaged chiefly in considering the mineral department of Geology; but it is clear that the Science could not have attained its present position without the continued labours of those who have been examining fossils in their relations to time and space. Had it not been for the researches of Palæontologists, Geology could not have made its present advance.
In my Class Book of Botany I have given an introduction to Palæophytology, and it occurred to me that it might be useful to students to publish this in a separate form, with additions in both the letterpress and the illustrations. The institution of the Palæontological Fellowship, in memory of my former friend Dr. Falconer, has brought the subject specially under my notice. The Fellowship has been promoted chiefly by my friend and former pupil Dr. Charles Murchison, a gentleman fond of science and of his Alma Mater, the University of Edinburgh, where he and Falconer studied and took their degrees.
The first award of the Fellowship has been made to a distinguished student, who acquitted himself with the greatest credit during the three days of examination on Geology,[ix] Zoology, and Botany. I trust that the Fellowship will continue to stimulate our eminent students in future years.
Having been a student of Natural Science along with Dr. Falconer, I feel a peculiar interest in doing what I can to promote the study of a subject to which he so successfully devoted his energies. In my endeavour to do so I have been encouraged by my friend and former pupil, Mr. William Carruthers, at the head of the Botanical Department of the British Museum, and a former student in Edinburgh under the late Professor Fleming. He has done much to advance our knowledge of Fossil Botany, and to him I am indebted for two of the plates and some of the woodcuts which illustrate this publication. He has given me most efficient assistance, and I have to return my best thanks for his kind aid. I am also indebted to my colleague, Professor Geikie, for his valued assistance.
The neighbourhood of Edinburgh is rich in Fossils of the Carboniferous epoch, and much yet remains to be done to illustrate its Palæontology. Such labourers as Geikie and Peach may be expected to give great assistance in the furtherance of our knowledge of Scottish Geology, so as to form a school which shall revive the reputation enjoyed by Edinburgh in the days of Hutton and Jameson. If I can be useful in encouraging students to take up the study of Palæontological Botany, and to prosecute it with vigour, I shall feel that this introductory treatise has not been issued in vain. As one of the few surviving relations of Dr. James Hutton, I am glad to be able to show an interest in a science which may aid in elucidating the "Theory of the Earth."
In writing this work I have taken for granted that the reader is acquainted with the Elements of Botany, and knows[x] the general structure of plants of the present day. I have not, therefore, hesitated to use the ordinary Botanical terms without explanation. I am satisfied that no one can study Fossil Botany properly unless he has studied Modern Botany.
Those readers who may find any difficulty as to technical terms I would refer to my Botanist's Companion, where a full Glossary is given.
27 Inverleith Row,
May 1872.
| PAGE | |
| Introductory Remarks | 1 |
| Determination of Fossil Plants | 3 |
| Mode of Preservation of Fossil Plants | 8 |
| Examination of the Structure of Fossil Plants | 12 |
| Fossiliferous Rocks | 20 |
| Natural Orders to which Fossil Plants belong | 22 |
| Periods of Vegetation among Fossil Plants | 25 |
| Fossil Flora of the Primary or Palæzoic Period | 26 |
| Reign of Acrogens | 26 |
| Flora of the Carboniferous Epoch | 36 |
| Flora of the Permian Epoch | 71 |
| Fossil Flora of the Secondary or Mesozoic Period | 72 |
| Reign of Gymnosperms | 72 |
| Flora of the Trias and Lias Epochs | 77 |
| Flora of the Oolitic Epoch | 80 |
| Flora of the Wealden Epoch | 84 |
| Fossil Flora of the Tertiary or Cainozoic Period (including the Cretaceous Epoch) | 87 |
| Reign of Angiosperms | 87 |
| Flora of the Chalk | 87 |
| [xii] Flora of the Eocene Epoch | 90 |
| Flora of the Miocene Epoch | 92 |
| Flora of the Pliocene Epoch | 98 |
| General Conclusions | 101 |
| Recapitulation | 103 |
| Works on Fossil Botany | 105 |
| Explanation of Plates | 111 |
| Index | 113 |
| Fig. | Page | |
| 1. | Section of Peuce Withami, Lindley and Hutton | 3 |
| 2. | Bark of Araucaria | 5 |
| 3. | Markings on Araucaria bark | 6 |
| 4. | "" | 7 |
| 5. | "" | 7 |
| 6. | Leaf of Araucaria | 7 |
| 7. | Nicolia Owenii (Carr.) | 11 |
| 8. | Bryson's instrument for slitting Fossils | 14 |
| 9. | Tree-fern | 27 |
| 10. | Asplenium | 28 |
| 11 | a. Bifurcating Trunk of a Tree-fern (Alsophila Perrottetiana) | 29 |
| 11 | b. Rhizome of Lastrea Filix-mas | 29 |
| 12. | Transverse section of stem of a Tree-fern (Cyathea) | 29 |
| 13. | Scalariform vessels from Tree-fern | 30 |
| 14. | Sporangia of a Fern | 30 |
| 15. | Lycopodium clavatum | 30 |
| 16. | Spore-case, containing Microspores of Lycopodium | 30 |
| 17. | ""Macrospores of Selaginella | 30 |
| 18. | Fructification of Equisetum maximum | 31 |
| 19. | Polygonal scale of Equisetum | 32 |
| 20. | Spore of Equisetum—filaments contracted | 32 |
| 21. | """expanded | 32 |
| 22. | Marsilea Fabri | 33 |
| 22 | bis. Adiantites Lindseæformis | 41 |
| 23. | Pecopteris (Alethopteris) aquiline | 43 |
| 24. | "(Alethopteris) heterophylla | 43 |
| 25. | Neuropteris Loshii | 43 |
| 26. | "gigantean | 43 |
| 27. | "acuminate | 43 |
| 28. | Sphenopteris affinis | 43 |
| 29. | Cyclopteris dilatata | 43 |
| [xiv] 30. | Stem of Caulopteris macrodiscus | 44 |
| 31. | ""Balfouri (Carr.) | 44 |
| 32. | ""Morrisi (Carr.) | 44 |
| 33. | "Sigillaria pachyderma | 45 |
| 34. | Sigillaria reniformis | 45 |
| 35. | "pachyderma | 46 |
| 36. | "(Favularia) tessellate | 46 |
| 37. | "pachyderma | 46 |
| 38. | Stigmaria ficoides | 47 |
| 39. | ""(S. anabathra of Corda) | 47 |
| 40. | Bifurcating stem of Lepidodendron obovatum (elegans) | 49 |
| 41. | Stem of Lepidodendron crenatum | 49 |
| 42. | Fructification of Lepidodendron | 50 |
| 43. | Longitudinal section of Fructification of Triplosporites | 50 |
| 44. | (1). Fruit of Selaginella spinulosa, A. Braun (Lycopodium selaginoides, Linn.) | 51 |
| (2). Scale and sporangium from the upper part of cone | 51 | |
| (3). Antheridian microspores from ditto | 51 | |
| (4). Macrospore | 51 | |
| (5). Scale and sporangium from lower part of cone, containing macrospores | 51 | |
| (6). Fruit of Lepidostrobus ornatus (Hooker) | 51 | |
| (7). Three scales and sporangia of ditto | 51 | |
| (8). Microspores from sporangia of the upper part of the cone of Triplosporites Brownii, Brongn. | 51 | |
| (9). Macrospore from the sporangia of the lower part | 51 | |
| (10). Scales and sporangia of a cone of Flemingites | 51 | |
| 45 | a. Calamites Suckovii | 57 |
| 45 | b. Septum or Phragma of a Calamite | 57 |
| 46. | Vertical stems of Calamites—in coal-measures of Treuil, near St. Etienne | 58 |
| 47. | Fruits of Equisetum and Calamites | 60 |
| (1). Equisetum arvense, L. | 60 | |
| (2). Portion of sporangium wall | 60 | |
| (3, 4). Spores—elaters free | 60 | |
| (5). Longitudinal section of part of one side of cone | 60 | |
| (6). Transverse section of cone | 60 | |
| (7). Calamites (Volkmannia) Binneyi (Carr.) | 60 | |
| (8). Portion of sporangium wall | 60 | |
| (9). Two spores | 60 | |
| [xv] (10). Longitudinal section of part of one side of cone | 60 | |
| (11). Transverse section of cone | 60 | |
| 48. | Foliage and fruits of Calamites | 62 |
| (1, 2). Asterophyllites | 62 | |
| (3, 4). Annularia | 62 | |
| (5, 6). Sphenophyllum | 62 | |
| 49. | Araucarioxylon Withami, Krauss (Pinites Withami) | 63 |
| 50. | Trigonocarpum olivæforme | 63 |
| 51. | Cardiocarpum Lindleyi (Carr.) | 65 |
| 52. | "" | 65 |
| 53. | Cardiocarpum anomalum (Carr.) | 66 |
| 54. | Pothocites Grantoni (Paterson) | 67 |
| 55, | 56. Walchia piniformis (Sternb.) | 72 |
| 57. | Pinus sylvestris | 73 |
| 58. | Abies excelsa | 73 |
| 59. | Larix Europæa | 73 |
| 60. | Cedrus Libani | 73 |
| 61. | Araucaria excelsa | 74 |
| 62. | Woody tubes of fir—single rows of discs | 74 |
| 63. | "" —double and opposite rows of discs | 74 |
| 64. | Woody tubes of Araucaria excelsa—double and triple and alternate rows of discs | 74 |
| 65. | Longitudinal section of stem of a Gymnosperm | 74 |
| 66. | Linear leaves of Pinus Strobus | 75 |
| 67. | Cone of Pinus sylvestris | 75 |
| 68. | "Cupressus sempervirens | 75 |
| 69. | Scale of mature cone of Pinus sylvestris | 75 |
| 70. | Fruiting branch of Juniperus communis | 76 |
| 71. | Branch of Taxus baccata | 76 |
| 72. | Male flower of Yew | 76 |
| 73. | Fruit of Yew | 76 |
| 74. | Cycas revoluta | 77 |
| 75. | Encephalartos (Zamia) pungens | 77 |
| 76. | Schizoneura heterophylla | 78 |
| 77. | Zamites | 79 |
| 78. | Pterophyllum Pleiningerii | 80 |
| 79. | Nilssonia compta (Pterophyllum comptum of Lindley and Hutton) | 80 |
| 80. | Palæozamia pectinata (Zamia pectinata of Brongniart, and Lindley and Hutton) | 80 |
| [xvi] 81. | Brachyphyllum mammillare | 81 |
| 82. | Equisetum columnare | 81 |
| 83. | Araucarites sphærocarpus (Carr.) | 82 |
| 84. | Termination of a scale of ditto | 82 |
| 85. | Section of a scale of ditto | 82 |
| 86. | The Dirt-bed of the island of Portland | 83 |
| 87. | Cycadoidea megalophylla (Mantellia nidiformis of Brongniart) | 83 |
| 88. | Kaidacarpum ooliticum (Carr.) | 84 |
| 89. | Pandanus odoratissimus | 84 |
| 90. | Fossil wood, Abietites Linkii | 85 |
| 91. | Sequoiites ovalis | 88 |
| 92. | Pinites ovatus (Zamia ovata of Lindley and Hutton) | 89 |
| 93. | Palmacites Lamanonis | 90 |
| 95. | Comptonia acutiloba | 92 |
| 96. | Acer trilobatum | 93 |
| 97. | Ulmus Bronnii | 93 |
| 98. | Rhamnus Aizoon | 94 |
| 99. | Alnus gracilis | 95 |
| 100. | Taxites or Taxodites Campbellii | 95 |
| 101. | Rhamnites multinervatus | 95 |
| 102. | Equisetum Campbellii | 96 |
The study of the changes which have taken place in the nature of living beings since their first appearance on the globe till the period when the surface of the earth, having assumed its present form, has been covered by the creation which now occupies it, constitutes one of the most important departments in Geology. It is, as Brongniart remarks, the history of life and its metamorphoses. The researches of geologists show clearly that the globe has undergone various alterations since that "beginning" when "God created the heavens and the earth." These alterations are exhibited in the different stratified rocks which form the outer crust of the earth, and which were chiefly sedimentary deposits produced by the weathering of the exposed rocks. Remains of the plants and animals living on the globe at the time of the formation of the different beds are preserved in them. Elevations and depressions of the surface of the earth affected the organisms on its surface, and gave to successive deposits new faunas and floras. Some of these epochs have been marked by great changes in the physical state of our planet, and they have been accompanied with equally great modifications in the nature of the living beings which inhabited it. The study of the fossil remains of animals is called Palæozoology (παλαιός, ancient, and ζῷον, animal), while the consideration of those of vegetables is denominated Palæophytology (παλαιός and φυτόν, a plant). Both are departments of the science of Palæontology, which[2] has been the means of bringing geology to its present state of advancement. The study of these extinct forms has afforded valuable indications as to the physical state of the earth, and as to its climate at different epochs. This study requires the conjunct labours of the Zoologist, the Botanist, and the Petralogist.
The vegetation of the globe, during the different stages of its formation, has undergone very evident changes. At the same time there is no reason to doubt that the plants may all be referred to the great classes distinguished at the present day—namely, Thallogens, including such plants as Lichens, Algæ, and Fungi; Acrogens, such as Ferns and Lycopods; Gymnosperms, such as Cone-bearing plants and Cycads; Endogens, such as Palms, Lilies, and Grasses; and Exogens, such as the common trees of Britain (excluding the Fir), and the great mass of ordinary flowering plants. The relative proportion of these classes, however, has been different, and the predominance of certain forms has given a character to the vegetation of different epochs. The farther we recede in geological history from the present day, the greater is the difference between the fossil plants and those which now occupy the surface. At the time when the coal-beds were formed, the plants covering the earth belonged to genera and species not existing at the present day. As we ascend higher, the similarity between the ancient and the modern flora increases, and in the latest stratified rocks we have in certain instances an identity in species and a considerable number of existing genera. At early epochs the flora appears to have been uniform, to have presented less diversity of forms than at present, and to have been similar in the different quarters of the globe. The vegetation also indicates that the nature of the climate was different from that which characterises the countries in which these early fossil plants are now found.
Fossil plants are by no means so easily examined as recent species. They are seldom found in a complete state. Fragments of stems, leaves, and fruits, are the data by which the plant is to be determined. It is very rare to find any traces of the flowers. The parts of fossil plants are usually separated from each other, and it is difficult to ascertain what are the portions which should be associated together so as to complete an individual plant. Specimens are sometimes preserved, so that the anatomical structure of the organs, especially of the stem, can be detected by very thin slices placed under the microscope. In the case of some stems the presence of punctated woody tissue (Fig. 1) has proved of great service as regards fossil Botany; this structure, along with the absence of large pitted ducts, serving to distinguish Conifers. The presence of scalariform vessels indicates a plant belonging to the vascular Cryptogams, of which the fern is the best known example. The cautions to be observed in determining fossil plants are noticed by Dr. Hooker in the Memoirs of the Geological Survey of Great Britain (vol. ii. p. 387). At the present day, the same fern may have different forms of fronds, which, unless they were found united, might be reckoned distinct genera; and remarkable examples are seen in Niphobolus rupestris and Lindsæa cordata. Moreover, we find the same form of frond belonging to several different genera, which can[4] only be distinguished by the fructification; and as this is rarely seen in fossil ferns, it is often impossible to come to a decided conclusion in regard to them. A leaf of Stangeria paradoxa was considered by an eminent botanist as a barren fern frond, but it ultimately proved to be the leaf of a Cycad. The leaf of Cupania filicifolia, a Dicotyledon, might easily be mistaken for that of a fern; it resembles much the frond of a fossil fern called Coniopteris. The diverse leaves of Sterculia diversifolia, if seen separately, might easily be referred to different plants. In the same fern we meet also with different kinds of venation in the fronds. Similar remarks may be made in regard to other plants. Harvey has pointed out many difficulties in regard to sea-weeds.
As regards the materials for a fossil flora, the following remarks of Hugh Miller deserve attention:—
"The authors of Fossil Floras, however able or accomplished they may be, have often to found their genera and species, and to frame their restorations, when they attempt these, on very inadequate specimens. For, were they to pause in their labours until better ones turned up, they would find the longest life greatly too short for the completion of even a small portion of their task. Much of their work must be of necessity of a provisional character—so much so, that there are few possessors of good collections who do not find themselves in circumstances to furnish both addenda and errata to our most valuable works on Palæontology. And it is only by the free communication of these addenda and errata that geologists will be at length enabled adequately to conceive of the by-past creations, and of that gorgeous Flora of the Carboniferous age, which seems to have been by far the most luxuriant and wonderful which our emphatically ancient earth ever saw."
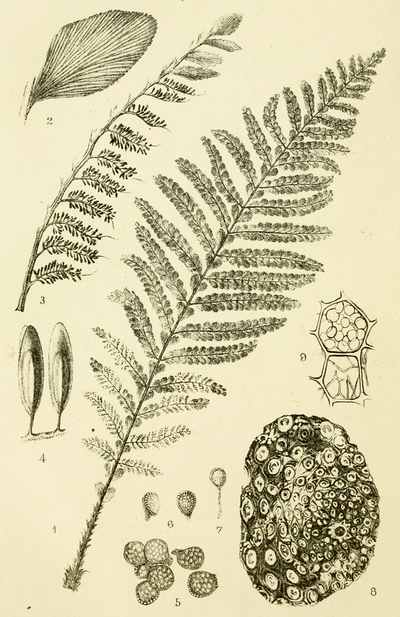
The bark of trees at the present day often exhibits different kinds of markings in its layers. This may be illustrated[5] by a specimen of Araucaria imbricata, which was destroyed by frost in the Edinburgh Botanic Garden on 24th December 1861. The tree was 24½ feet high, with a circumference of four feet at the base of the stem, and had twenty whorls of branches. The external surface of the bark is represented in Fig. 2. There are seen scars formed in part by prolongations from the lower part of the leaves, which have been cut off close to their union with the stem. The base of each leaf remaining in the bark has the form of a narrow elongated ellipse, surrounded by cortical foliar prolongations. The markings on the bark, when viewed externally, have a somewhat oblique quadrilateral form. On removing the epiphlœum or outer bark, and examining its inner surface, we remark a difference in the appearance presented at the lower and upper part of the stem. In the lower portion the markings have an irregular elliptical form, with a deep depression, and fissures where the leaves are attached (Fig. 3). Higher up the epiphlœal markings assume rather more of a quadrilateral form, with the depressions less deep, and the fissures for the leaves giving off prolongations on[6] either side. Farther up the markings are smaller in size, obliquely quadrilateral, and present circular clots along the boundary lines chiefly (Fig. 4). Higher still the quadrilateral form becomes more apparent, and the dots disappear (Fig. 5). The epiphlœum thus presents differences in its markings at different heights on the stem.
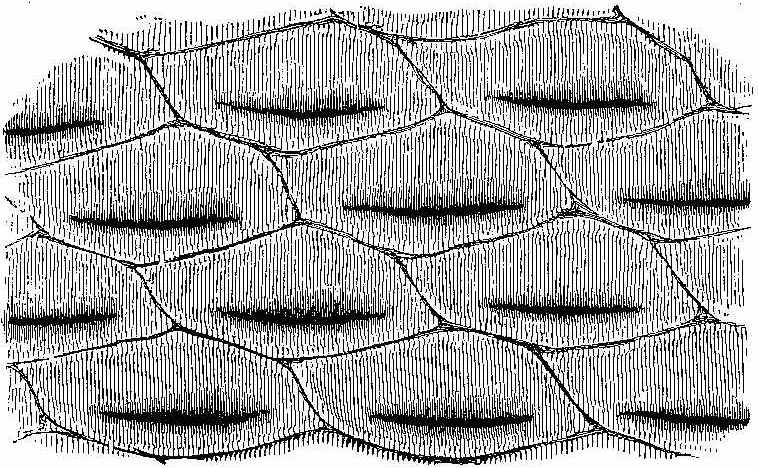


The part of the bark immediately below the epiphlœum is well developed, and is of a spongy consistence. When examined microscopically it is seen to be composed of cells of[7] various shapes—some elongated fusiform, others rhomboidal, others with pointed appendages. The variety of forms is very great, but it is possible that this may be partly owing to the effects of frost on the cells. On the spontaneous separation of the bark, the portion below the epiphlœum was seen to consist of distinct plates of a more or less quadrilateral form, with some of the edges concave and others convex, a part in the centre indicating the connection with the leaf, along with which it is detached. In Fig. 6 a leaf is shown with one of these plates attached.
The appearances presented by the outer and middle bark of Araucaria imbricata bear a marked resemblance to those exhibited by certain fossils included in the genera Sigillaria and Lepidodendron. The sculpturesque markings on the stems of these fossil plants indicate their alliance to the ferns and lycopods of the present epoch. But it is evident, from these markings, that much caution is required in making this determination. Other points of structure must be examined before a proper decision can be formed. When,[8] for instance, the presence of scalariform tissue, or of punctated woody tissue, has been satisfactorily shown under the microscope, we are entitled to hazard an opinion as to the affinities of the fossils. In many instances, however, external appearances are the only data on which to rely for the determination of fossil genera and species; and rash conclusions have often been drawn by geologists who have not been conversant with the structure of plants. The Araucaria markings point out the need of care in drawing conclusions, and their variations at different parts of the bark indicate the danger of a rash decision as to species. There can be no doubt that in vegetable Palæontology the number of species has been needlessly multiplied—any slight variation in form having been reckoned sufficient for specific distinction. We can conceive that the Araucaria bark markings in a fossil state might easily supply several species of Lepidodendron. A naturalist, with little knowledge of the present flora of the globe, ventures sometimes to decide on an isolated fragment. Hence the crude descriptions of fossil vegetable forms, and the confusion in which Palæophytology is involved. Every geologist who examines fossil plants ought to be well acquainted with the minute structure of living plants, the forms of their roots, stems, leaves, fronds, and fructification; the markings on the outer and inner surfaces of their barks, on their stems, and on their rhizomes; the localities in which they grow, and the climates which genera and species affect in various parts of the world. (Professor Balfour in the Proceedings of the Royal Society of Edinburgh, April 1862, vol. iv. p. 577.)
The mode in which plants are preserved in a fossil state may be referred to four principal classes:—1. Casts of the plants; from which all the original substance and structure have been removed subsequently to the burial of the plants, and to the[9] greater or less induration of the rocks in which they are entombed. Such casts are occasionally hollow, but more frequently they consist of the amorphous substance of the rock which has filled up the cavity, and which exhibits, often with remarkable minuteness, the external aspects of the original specimen. 2. Carbonisation; in which the original substance of the plant has been chemically altered and converted into lignite or coal. All trace of the form of the original plant is generally lost, as is the case with the extensive beds of coal; but frequently, when the organism has been buried in a bed of clay, the external appearance is faithfully preserved, as in the ferns and other foliage found in the shales of the coal-measures. 3. Infiltration; in which the vegetable tissues, though carbonised, retain their original form from the infiltration of some mineral in solution, chiefly lime or silex, which has filled the empty cells and vessels, and so preserved their original form. This mode of preservation occurs in the calcareous nodules in coal-beds, in the remarkable ash-beds discovered by Mr. Wünsch in Arran, and generally in the secondary rocks. 4. Petrifaction; in which the structure is preserved, but the whole of the original substance has been replaced, atom for atom, by an inorganic substance, generally lime, silex, or some ore of iron. This is the condition of the beautiful fossils from Antigua, and of many stems and fruits from rocks of all ages in Britain.
Carbonised vegetables, or those which have passed into the state of Lignites, often undergo modifications which render it difficult to understand them rightly. Sometimes a portion of the organs of vegetables which have passed into the state of lignite is transformed into pyrites, or else pyrites of a globular shape is found in the middle of the tissue, and may be taken for a character of organisation. The section of certain Dicotyledonous fossil woods, in that case, may resemble Monocotyledons. Petrifaction, as in the case of silicified[10] woods, often preserves all the tissues equally, at other times the soft tissues are altered or destroyed; the cellular tissue being replaced by amorphous chalcedony, while the ligneous and vascular tissues alone are petrified, so as to preserve their forms. In some cases the reverse takes place as to these tissues; the fibrous portions disappear, leaving cavities, while the cells are silicified. Sometimes we find the parts regularly silicified at one place, so as to retain the structure, while at another an amorphous mass of silica is found. In such cases there appear, as it were, distinct silicified woody bundles in the midst of an amorphous mass. The appearance depends, however, merely on irregular silicification or partial petrifaction. Infiltrated fossil woods, by means of chemical tests, are shown to possess portions of vegetable tissues cemented into a mass by silica. In some cases we find the vessels and cells separately silicified, without being crushed into a compact mass. In these cases, the intercellular substance not being silicified, the mass breaks down easily; whereas, when complete silicification takes place, the mass is not friable. Coniferous wood is often friable, from silicified portions being still separated from each other by vegetable tissue more or less entire. During silicification, or subsequent to it, it frequently happens that the plant has been compressed, broken, and deformed, and that fissures have been formed which have been subsequently filled with crystallised or amorphous silica.
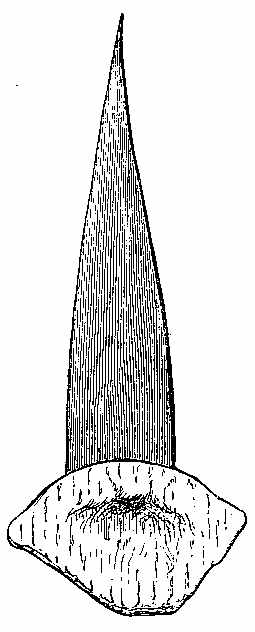
Silicified stems of trees have been observed in various parts of the world, with their structure well preserved, so that their Endogenous and Exogenous character could be easily determined. The Rev. W. B. Clarke notices the occurrence of a fossil pine-forest at Kurrur-Kurrân, in the inlet of Awaaba, on the eastern coast of Australia. In the inlet there is a formation of conglomerate and sandstone, with subordinate beds of lignite—the lignite forming the so-called Australian coal. Throughout the alluvial flat, stumps and[11] stools of fossilised trees are seen standing out of the ground, and one can form no better notion of their aspect than by imagining what the appearance of the existing living forest of Eucalypti and Casuarinæ would be if the trees were all cut down to a certain level. In a lake in the vicinity there are also some fossilised stumps of trees, standing vertically. In Derwent Valley, Van Diemen's Land, fossil silicified trees, in connection with trap rocks, have been found in an erect position. One was measured with a stem 6 feet high, a circumference at the base of 7 feet 3 inches, and a diameter at the top of 15 inches. The stems are Coniferous, resembling Araucaria. The outer portion of the stem is of a rich brown glossy agate, while the interior is of a snowy whiteness. One hundred concentric rings have been counted. The tissue falls into a powdery mass. Silica is found in the inside of the tubes, and their substance is also silicified. The erect silicified stems of coniferous trees exist in their natural positions in the "dirt-bed," an old surface soil in the sandstone strata of the Purbeck series in the Isle of Portland, Dorsetshire. In the petrified forests near Cairo silicified stems have been examined by Brown, Unger, and Carruthers. They belong to dicotyledonous trees (not coniferous), to which the names of Nicolia Ægyptiaca and Nicolia Owenii (Fig. 7) have been given. The wood consists of a slender prosenchyma, abundantly penetrated by large ducts. The walls of the ducts are marked by small, regularly arranged, oval, and somewhat compressed hexagonal reticulations. The[12] ducts have transverse diaphragms. There are numerous medullary rays. The wood in their stems is converted into chalcedony. (Carruthers on Petrified Forest near Cairo. Geol. Mag., July 1870.)
When the structure of fossil plants is well preserved, it may be seen under the microscope by making thin sections after the mode recommended by Mr. William Nicol, the inventor of the prism which bears his name, and to whose memory Unger dedicated the genus Nicolia, which has just been described as constituting the petrified forest at Cairo. The following is a description of the process of preparing fossils for the microscope, by Mr. Alexander Bryson. (Edin. N. Phil. Journal, N. S. iii. 297. Balfour's Botanist's Companion, p. 30.)
"The usual mode of proceeding in making a section of fossil wood is simple, though tedious. The first process is to flatten the specimen to be operated on by grinding it on a flat lap made of lead charged with emery or corundum powder. It must now be rendered perfectly flat by hand on a plate of metal or glass, using much finer emery than in the first operation of grinding. The next operation is to cement the object to the glass plate. Both the plate of glass and the fossil to be cemented must be heated to a temperature rather inconvenient for the fingers to bear. By this means moisture and adherent air are driven off, especially from the object to be operated on. Canada balsam is now to be equally spread over both plate and object, and exposed again to heat, until the redundant turpentine in the balsam has been driven off by evaporation. The two surfaces are now to be connected while hot, and a slow circular motion, with pressure, given either to the plate or object, for the purpose of throwing out the superabundant balsam and globules of[13] included air. The object should be below and the glass plate above, as we then can see when all the air is removed, by the pressure and motion indicated. It is proper to mention that too much balsam is more favourable for the expulsion of the air-bubbles than too little. When cold, the Canada balsam will be found hard and adhering, and the specimen fit for slitting. This process has hitherto been performed by using a disc of thin sheet-iron, so much employed by the tinsmith, technically called sheet-tin. The tin coating ought to be partially removed by heating the plate, and when hot rubbing off much of the extraneous tin by a piece of cloth. The plate has now to be planished on the polished stake of the tinsmith, until quite flat. If the plate is to be used in the lathe, and by the usual method, it ought to be planished so as to possess a slight convexity. This gives a certain amount of rigidity to the edge, which is useful in slitting by the hand; while by the method of mechanical slitting, about to be described, this convexity is inadmissible. The tin plate, when mounted on an appropriate chuck in the lathe, must be turned quite true, with its edge slightly rounded and made perfectly smooth by a fine-cut file. The edge of the disc is now to be charged with diamond powder. This is done by mingling the diamond powder with oil, and placing it on a piece of the hardest agate, and then turning the disc slowly round. Then, by holding the agate with the diamond powder with a moderate pressure against the edge of the disc, it is thoroughly charged with a host of diamond points, becoming, as it were, a saw with invisible teeth. In pounding the diamond, some care is necessary, as also a fitting mortar. The mortar should be made of an old steel die, if accessible; if not, a mass of steel, slightly conical, the base of which ought to be 2 inches in diameter, and the upper part 1½ inch. A cylindrical hole is now to be turned out in the centre, of ¾ths of an inch diameter, and about 1 inch deep. This, when hardened, is the mortar;[14] for safety it may be annealed to a straw colour. The pestle is merely a cylinder of steel, fitting the hollow mortar but loosely, and having a ledge or edging of an eighth of an inch projecting round it, but sufficiently raised above the upper surface of the mortar, so as not to come in contact while pounding the diamond. The point of the pestle ought only to be hardened and annealed to a straw colour, and should be of course convex, fitting the opposing and equal concavity of the mortar. The purpose of the projecting ledge is to prevent the smaller particles of diamond spurting out when the pestle is struck by the hammer."
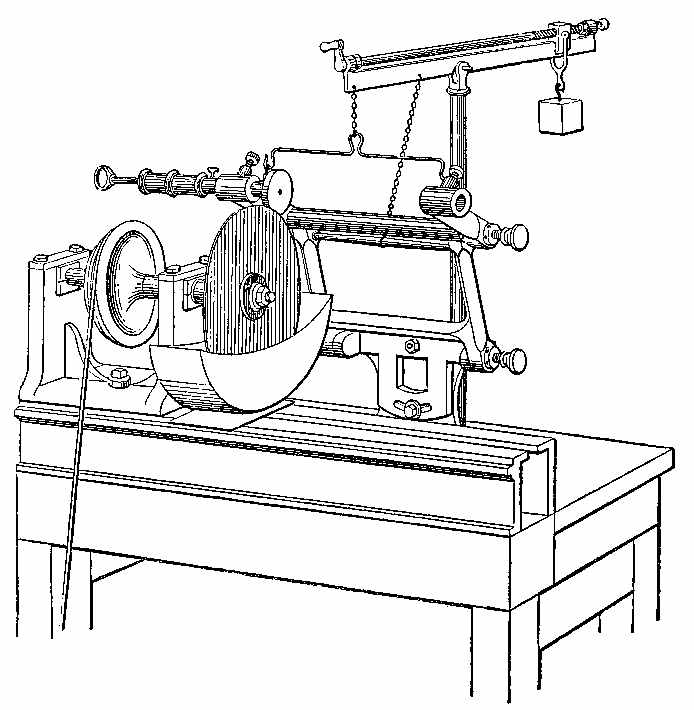
Mr. Bryson has contrived an instrument for slitting fossils. The instrument is placed on the table of a common lathe, which is, of course, the source of motion (Fig. 8). It consists[15] of a Watt's parallel motion, with four joints, attached to a basement fixed to the table of the lathe. This base has a motion (for adjustment only) in a horizontal plane, by which we may be enabled to place the upper joint in a parallel plane with the spindle of the lathe. This may be called the azimuthal adjustment. The adjustment, which in an astronomical instrument is called the plane of right ascension, is given by a pivot in the top of the base, and clamped by a screw below. This motion in right ascension gives us the power of adjusting the perpendicular planes of motion, so that the object to be slit passes down from the circumference of the slitting-plate to nearly its centre, in a perfectly parallel plane. When this adjustment is made accurately, and the slitting-plate well primed and flat, a very thin and parallel slice is obtained. This jointed frame is counterpoised and supported by a lever, the centre of which is movable in a pillar standing perpendicularly from the lathe table. Attached to the lever is a screw of three threads, by which the counterpoise weight is adjusted readily to the varying weight of the object to be slit and the necessary pressure required on the edge of the slitting-plate.
The object is fixed to the machine by a pneumatic chuck. It consists of an iron tube, which passes through an aperture on the upper joint of the guiding-frame, into which is screwed a round piece of gun-metal, slightly hollowed in the centre, but flat towards the edge. This gun-metal disc is perforated by a small hole communicating with the interior of the iron tube. This aperture permits the air between the glass plate and the chuck to be exhausted by a small air-syringe at the other end. The face of this chuck is covered with a thin film of soft india-rubber not vulcanised, also perforated with a small central aperture. When the chuck is properly adjusted, and the india-rubber carefully stretched over the face of the gun-metal, one or two pulls of the syringe-piston is quite[16] sufficient to maintain a very large object under the action of the slitting-plate. By this method no time is lost; the adhesion is made instantaneously, and as quickly broken by opening a small screw, to admit air between the glass plate and the chuck, when the object is immediately released. Care must be taken, in stretching the india-rubber over the face of the chuck, to make it very equal in its distribution, and as thin as is consistent with strength. When this material is obtained from the shops, it presents a series of slight grooves, and is rather hard for our purpose. It ought, therefore, to be slightly heated, which renders it soft and pliant, and in this state should now be stretched over the chuck, and a piece of soft copper wire tied round it, a slight groove being cut in the periphery of the chuck to detain the wire in its place. When by use the surface of the india-rubber becomes flat, smooth, and free from the grooves which at first mar its usefulness, a specimen may be slit of many square inches, without resort being had to another exhaustion by the syringe. But when a large, hard, siliceous object has to be slit, it is well for the sake of safety to try the syringe piston, and observe if it returns forcibly to the bottom of the cylinder, which evidences the good condition of the vacuum of the chuck.
After the operation of slitting, the plate must be removed from the spindle of the lathe, and the flat lead lap substituted. The pneumatic chuck is now to be reversed, and the specimen placed in contact with the grinder. By giving a slightly tortuous motion to the specimen, that is, using the motion of the various joints, the object is ground perfectly flat when the length of both arms of the joints is perfectly equal. Should the leg of the first joint on the right-hand side be the longer, the specimen will be ground hollow; if shorter, it will be ground convex. But if, as before stated, they are of equal length, a perfectly parallel surface will be obtained.
In operating on siliceous objects, I have found soap and water quite as speedy and efficacious as oil, which is generally used; while calcareous fossils must be slit by a solution of common soda in water. This solution of soda, if made too strong, softens the india-rubber on the face of the pneumatic chuck, and renders a new piece necessary; but if care is taken to keep the solution of moderate strength, one piece of india-rubber may last for six months. The thinner and flatter it becomes, the better hold the glass takes, until a puncture occurs in the outer portion, and a new piece is rendered necessary.
The polishing of the section is the last operation. This is performed in various ways, according to the material of which the organism is composed. If siliceous, a lap of tin is to be used, about the same size as the grinding lap. Having turned the face smooth and flat, a series of very fine notches are to be made all over the surface. This operation is accomplished by holding the edge of an old dinner-knife almost perpendicular to the surface of the lap while rotating; this produces a series of criddles, or slight asperities, which detain the polishing substance. The polishing substance used on the tin lap is technically called lapidaries' rot-stone, and is applied by slightly moistening the mass, and pressing it firmly against the polisher, care being taken to scrape off the outer surface, which often contains grit. The specimen is then to be pressed with some degree of force against the revolving tin lap or polisher, carefully changing the plane of action, by moving the specimen in various directions over the surface.
To polish calcareous objects, another method must be adopted as follows:—
A lap or disc of willow wood is to be adapted to the spindle of the lathe, three inches in thickness, and about the diameter of the other laps (10 inches), the axis of the wood being parallel to the spindle of the lathe, that is, the acting[18] surface of the wood is the end of the fibres, the section being transverse.
This polisher must be turned quite flat and smoothed by a plane, as the willow, from its softness, is peculiarly difficult to turn. It is also of consequence to remark that both sides should be turned, so that the lap, when dry, is quite parallel. This lap is most conveniently adapted to the common face chuck of a lathe with a conical screw, so that either surface may be used. This is made evident, when we state that this polisher is always used moist, and, to keep both surfaces parallel, must be entirely plunged in water before using, as both surfaces must be equally moist, otherwise the dry surface will be concave and the moist one convex. The polishing substance used with this lap is putty powder (oxide of tin), which ought to be well washed, to free it from grit. The calcareous fossils being finely ground, are speedily polished by this method. To polish softer substances, a piece of cloth may be spread over the wooden lap, and finely-levigated chalk used as a polishing medium.
In order to study fossil plants well, there must be an acquaintance with systematic botany, a knowledge of the microscopical structure of all the organs of plants, such as their roots, stems, barks, leaves, fronds, and fruit; of the markings which they exhibit on their different surfaces, and of the scars which some of them leave when they decay. It is only thus we can expect to determine accurately the living affinities of the fossil. Brongniart says, that before comparing a fossil vegetable with living plants, it is necessary to reconstruct as completely as possible the portion of the plant under examination, to determine the relations of these portions to the other organs of the same plant, and to complete the plant if possible, by seeing whether, in the fossils of the same locality, there may not be some which belong to the same plant. The connection of the different parts of the[19] same plant is one of the most important problems in Palæophytology, and the neglect of it has led to many mistakes. In some instances the data have been sufficient to enable botanists to refer a fossil plant to a genus of the present day, so that we have fossil species of the genera Ulmus, Alnus, Pinus, etc. Sometimes the plant is shown to be allied to a living genus, but differing in some essential point, or wanting something to complete the identity, and it is then marked by the addition of the term ites, as Pinites, Thuites, Zamites, etc.
Before drawing conclusions as to the climate or physical condition of the globe at different geological epochs, the botanist must be well informed as to the vegetation of different countries, as to the soils and localities in which certain plants grow, whether on land or in the sea, or in lakes, in dry or marshy ground, in valleys or on mountains, or in estuaries, in hot, temperate, or cold regions. Great caution must be employed also in predicating from one species the conditions of another, inasmuch as different species of the same genus frequently exist in very different habitats, and under almost opposite conditions of moisture and temperature. It is only by a careful consideration of all these particulars that any probable inferences can be drawn as to the condition of the globe. Considering the physiognomy of vegetation at the present day, we find remarkable associations of forms. The Palms, although generally characteristic of very warm countries, are by no means confined to them; Chamærops humilis extending to Europe as far as lat. 43° to 44° N., and C. palmetto in North America to lat. 34° to 36° N., while C. Fortunei, from the north of China, is perfectly hardy in the south of England. Major Madden mentions the association of Palms and Bamboos with Conifers at considerable elevations on the Himalayas. (Edin. Bot. Soc. Trans. iv., p. 185.) Epiphytic Orchids, which usually characterise[20] warm climates, have representatives at great elevations, as Oncidium nubigenum at 14,000 feet in the Andes, and Epidendrum frigidum at from 12,000 to 13,000 feet in the Columbia mountains. These facts point out the care necessary before drawing conclusions as to the climate which fossil plants may be supposed to indicate.
The rocks of which the globe is composed are divided into two great classes—the Stratified or Aqueous, and the Unstratified or Igneous. The stratified rocks frequently contain fossil remains, and are then called fossiliferous; those with no such remains are designated non-fossiliferous or azoic. The igneous unstratified rocks, included under the names of Granitic and Trappean, show no appearance of animal or vegetable remains. Those trap rocks, however, which have been formed of loose volcanic ashes have often enclosed and preserved the remains of plants and animals; while even between the successive beds of old lava-like trap rocks organic remains are sometimes found. Thus, in Antrim, near the Giant's Causeway, deposits containing vegetable remains occur inter-stratified with basaltic rocks. These remains are of Miocene age, and have been referred to coniferous plants, beeches, oaks, plane trees, etc. Similar plants have been discovered in a similar position by the Duke of Argyll in the island of Mull. In trap rocks near Edinburgh, lignite with distinct structure has also been detected. Silicified wood and coal, imbedded in trap rocks, have been seen in Kerguelen's Land. The wood is found enclosed in basalt, whilst the coal crops out in ravines, in close contact with the overlying porphyritic and amygdaloidal greenstone. Hooker has also seen silicified wood, in connection with trap, in Macquarrie's Plains, in Tasmania. Several beds of trap-tuff or ash, formed into[21] solid compact rock by infiltrated carbonate of lime, occur in the north-east of Arran, which contain numerous stems, branches, and fruits of carboniferous plants. These represent the remains of successive forests which grew on this locality, and were one after the other destroyed by the ash-showers poured forth from a neighbouring volcano during its intermittent periods of activity.
Fossil remains are extremely rare in certain rocks, which, from the changes they have undergone, have been denominated Metamorphic. These include Gneiss and Mica-slate, which are stratified rocks subsequently altered by heat and other causes, and so completely metamorphosed that the traces of organisms have been nearly obliterated. Nevertheless, recognisable traces of plant and animal remains have been found in what were recently thought to be azoic rocks. The absence of organic remains in rocks is therefore not sufficient to enable us to state that these rocks were formed before animals or vegetables existed.
The stratified rocks which contain fossils have been divided into three great groups—the Palæozoic, the Secondary, and the Tertiary, or into Palæozoic and Neozoic groups. The formations included under these are exhibited in the following table, taken from Lyell's Manual of Geology:—
| 1. | Recent. | } | Post Tertiary. | } | Recent. | ||
| 2. | Post Pliocene. | } | } | ||||
| 3. | Newer Pliocene. | } | Pliocene. | } | Tertiary or Cainozoic. | } | |
| 4. | Older Pliocene. | } | } | } | |||
| } | } | ||||||
| 5. | Upper Miocene. | } | Miocene. | } | } | ||
| 6. | Lower Miocene. | } | } | } | |||
| } | } | ||||||
| 7. | Upper Eocene. | } | Eocene. | } | } | ||
| 8. | Middle Eocene. | } | } | } | |||
| 9. | Lower Eocene. | } | } | } | Neozoic. | ||
| } | |||||||
| 10. | Maestricht Beds. | } | } | } | |||
| 11. | White Chalk. | } | } | } | |||
| 12 | Chloritic Series. | } | } | Secondary | } | ||
| 13. | Gault | } | Cretaceous. | } | or | } | |
| 14. | Neocomian. | } | } | Mesozoic. | } | ||
| 15. | Wealden. | } | } | } | |||
| [22] 16. | Purbeck Beds. | } | } | } | |||
| 17. | Portland Stone. | } | } | } | |||
| 18. | Kimmeridge Clay. | } | } | } | |||
| 19. | Coral Rag. | } | Jurassic. | } | } | ||
| 20. | Oxford Clay. | } | } | Secondary | } | ||
| 21. | Great or Bath Oolite. | } | } | or | } | Neozoic. | |
| 22. | Inferior Oolite. | } | } | Mesozoic. | } | ||
| 23. | Lias. | } | } | } | |||
| } | } | ||||||
| 24. | Upper Trias. | } | } | } | |||
| 25. | Middle Trias. | } | Triassic. | } | } | ||
| 26. | Lower Trias. | } | } | } | |||
| 27. | Permian. | Permian. | } | ||||
| } | |||||||
| 28. | Coal Measures. | } | } | ||||
| 29. | Carboniferous | } | Carboniferous. | } | |||
| limestone. | } | } | |||||
| } | |||||||
| 30. | Upper Devonian. | } | Devonian | } | |||
| 31. | Middle Devonian. | } | or Old Red | } | |||
| 32. | Lower Devonian. | } | Sandstone. | } | Primary | } | |
| } | } | ||||||
| 33. | Upper Silurian. | } | Silurian. | } | or | } | Palæozoic. |
| 34. | Lower Silurian. | } | } | } | |||
| } | Palæozoic. | } | |||||
| 35. | Upper Cambrian. | } | Cambrian. | } | |||
| 36. | Lower Cambrian. | } | } | ||||
| } | |||||||
| 37. | Upper Laurentian. | } | Laurentian. | } | |||
| 38. | Lower Laurentian. | } | } | ||||
The plants found in different strata are either terrestrial or aquatic, and the latter exhibit species allied to the salt and fresh water vegetables of the present day. Their state of preservation depends much on their structure. Cellular plants have probably in a great measure been destroyed, and hence their rarity; while those having a woody structure have been preserved. The following is the number of fossil genera and species, as compiled from Unger's work on Palæophytology—(Unger, Genera et Species Plantarum Fossilium, 1850).
| Dicotyledones. | Genera. | Species. | |
| Thalamifloræ. | 24 | 84 | |
| Calycifloræ | 56 | 182 | |
| Corollifloræ | 23 | 60 | |
| Monochlamydeæ Angiospermæ | 48 | 221 | |
| —————— Gymnospermæ | 56 | 363 | |
| [23]Monocotyledones. | |||
| Petaloideæ | 38 | 130 | |
| Glumiferæ | 5 | 12 | |
| Acotyledones. | |||
| Thallogenæ | 31 | 203 | |
| Acrogenæ | 121 | 969 | |
| Doubtful | 35 | 197 | |
| —— | —— | ||
| 437 | 2421 | ||
These plants are arranged in the different strata as follows:—
| { | Cambrian, Silurian, and Devonian | 73 | |
| Palæozoic | { | Carboniferous | 683 |
| { | Permian | 97 | |
| { | Triassic | 115 | |
| Mesozoic | { | Jurassic | 294 |
| { | Cretaceous | 183 | |
| { | Eocene | 414 | |
| Cainozoic | { | Miocene | 496 |
| { | Pliocene | 35 | |
| Recent | Post-Pliocene | 31 | |
| —— | |||
| Fossil Species. | 2421 |
During the twenty years that have elapsed since this enumeration was made, the number of fossil species has been very greatly increased. The proportion exhibited in this table is likewise greatly altered from the enormous additions made to the Tertiary Flora by Unger, Ettingshausen, and Heer, and from the important contributions by Principal Dawson to the Devonian Flora.
Among the fossil Thalamifloral Dicotyledons, Unger mentions species belonging to the orders—
Among Calycifloral Dicotyledons—
Among Corollifloral Dicotyledons—
Among Monochlamydeous Angiosperms—
Among Monochlamydeous Gymnosperms—
Among Petaloid Monocotyledons—
Among Glumiferous Monocotyledons—
Among Acrogenous Acotyledons—
Among Thallogenous Acotyledons—
On taking a general survey of the known fossil plants, Brongniart thought that he could trace three periods of vegetation, characterised by the predominance of certain marked forms of plants. In the ancient period there is a predominance of Acrogenous Cryptogamic plants; this is succeeded by a period in which there is a preponderance of Gymnospermous Dicotyledons; while a third period is marked by the predominance of Angiospermous Dicotyledons. There is thus—1. The reign of Acrogens, which includes the plants of the Devonian, Carboniferous, and Permian periods. During these periods there seems to be a predominance of Ferns, and a great development of arborescent Lycopodiaceæ, such as Lepidodendron and Sigillaria, and with them are associated some Gynmosperms, allied to Araucaria, and some anomalous plants, as Noeggerathia. 2. The reign of Gymnosperms, comprehending the Triassic and Jurassic periods. Here we meet with numerous Coniferæ and Cycadaceæ, while Ferns are less abundant. 3. The reign of Angiosperms, embracing the Cretaceous and the Tertiary periods. This is characterised by the predominance of Angiospermous Dicotyledons, a class of plants which constitute more than three-fourths of the present vegetable productions of the globe, and which appear to have acquired a predominance from the commencement of the Tertiary formations. These plants appear sparingly even at the beginning of the chalk formation in Europe, but are more abundant in this formation as developed in North America.
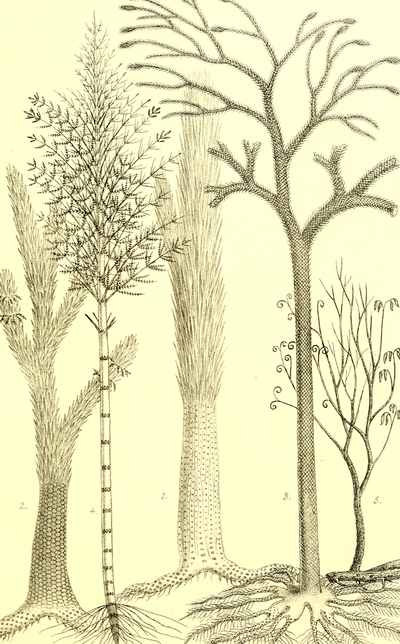
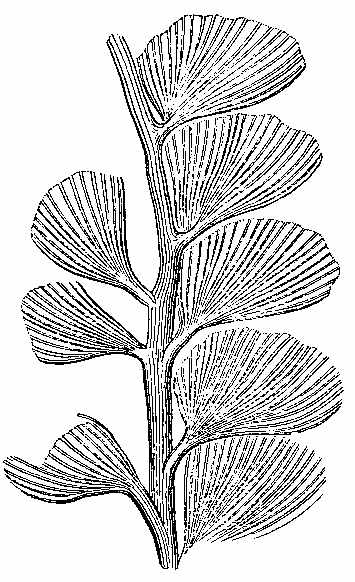
In the present day, acrogenous plants are represented by cellular and vascular Cryptogams. In considering fossil plants our attention is specially directed to the latter. In the recent Floras, vascular Acrogens are represented by such plants as Ferns, Lycopods, and Equisetums. Some of them have an arborescent habit, but the greater number are shrubby and herbaceous. Many of them have creeping rhizomes, which are either subterranean, or run along the surface of the ground. One of these arborescent forms is seen in Tree-ferns (Fig. 9). Another form with a rhizome is seen in Fig. 10. The trunks of ferns are marked by scars, which indicate the parts where the bases of the fronds were attached, and where the vascular tissue passes out from the interior (Fig. 11, a and b). A transverse section of the stem (Fig. 12) shows a continuous cylinder of scalariform vessels (Fig. 13), enclosing a large mass of cellular tissue frequently penetrated by small scalariform bundles. The cylinder is pierced by meshes, from the inner sides of which rise the vascular bundles going to the leaves, while some of the free bundles of the axis pass through the mesh, carrying with them a portion of the cellular tissue into the petiole. The fructification consists of spore-cases (sporangia), often with an elastic ring round them, containing spores in their interior (Fig. 14).

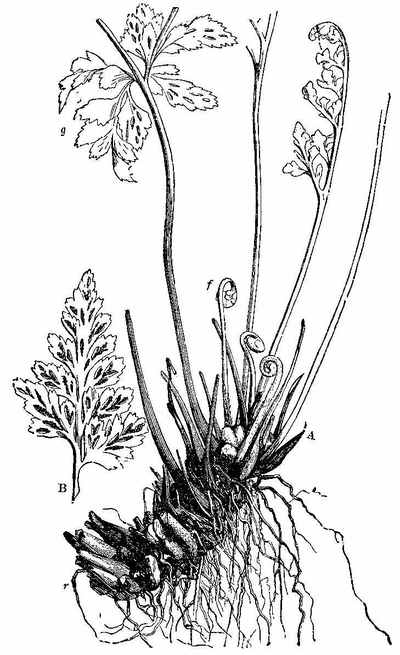
Among Acrogens of the present day there are also plants belonging to the natural order Lycopodiaceæ or Club-mosses (Fig. 15), having creeping stems, which give rise to leafy branches. The leaves are small, sessile, and moss-like, and the fructification consists of two kinds of cellular bodies,[27] small spores or microspores (Fig. 16), and large spores or macrospores (Fig. 17). They consist of cellular and vascular tissues, the latter occurring in the form of woody, annular, and scalariform vessels, which occupy the axis or central part of the stem. They differ from ferns in the distribution of their vascular bundles. The order is represented also by such plants as Selaginella, Psilotum, Phylloglossum, and Isoetes. In the plant called Isoetes (Quillwort) there is a peculiar short stem[28] which does not increase in height. It produces additions laterally, so that the stem increases in thickness. The leaves[29] continue to multiply, and bear fructification at their bases. They have both large and small spores.

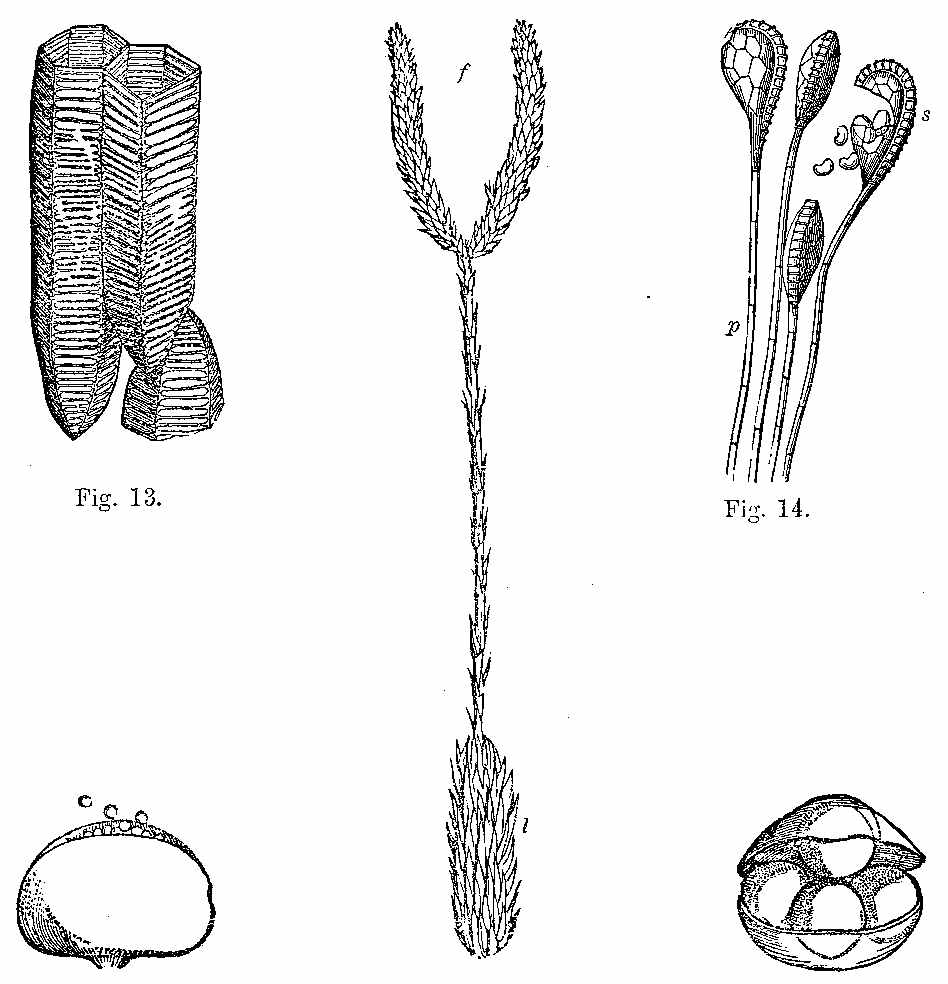

Another important order of vascular Acrogens is the Equisetaceæ or Horse-tails (Fig. 18). These are Cryptogams, having rhizomes, bearing hollow, striated branches, which secrete in their epidermis a considerable amount of silex. These branches are jointed and have membranous sheaths at the articulations, which are whorls of leaves reduced to a very rudimentary condition. The fructification consists of cone-like bodies (Fig. 18, f) bearing peltate polygonal scales, under which are spore-cases (Fig. 19), enclosing spores with[30] four hygrometric club-shaped filaments called elaters (Figs. 20 and 21). At the present day some of these plants in tropical regions[31] have stems of 15 or 16 feet high.
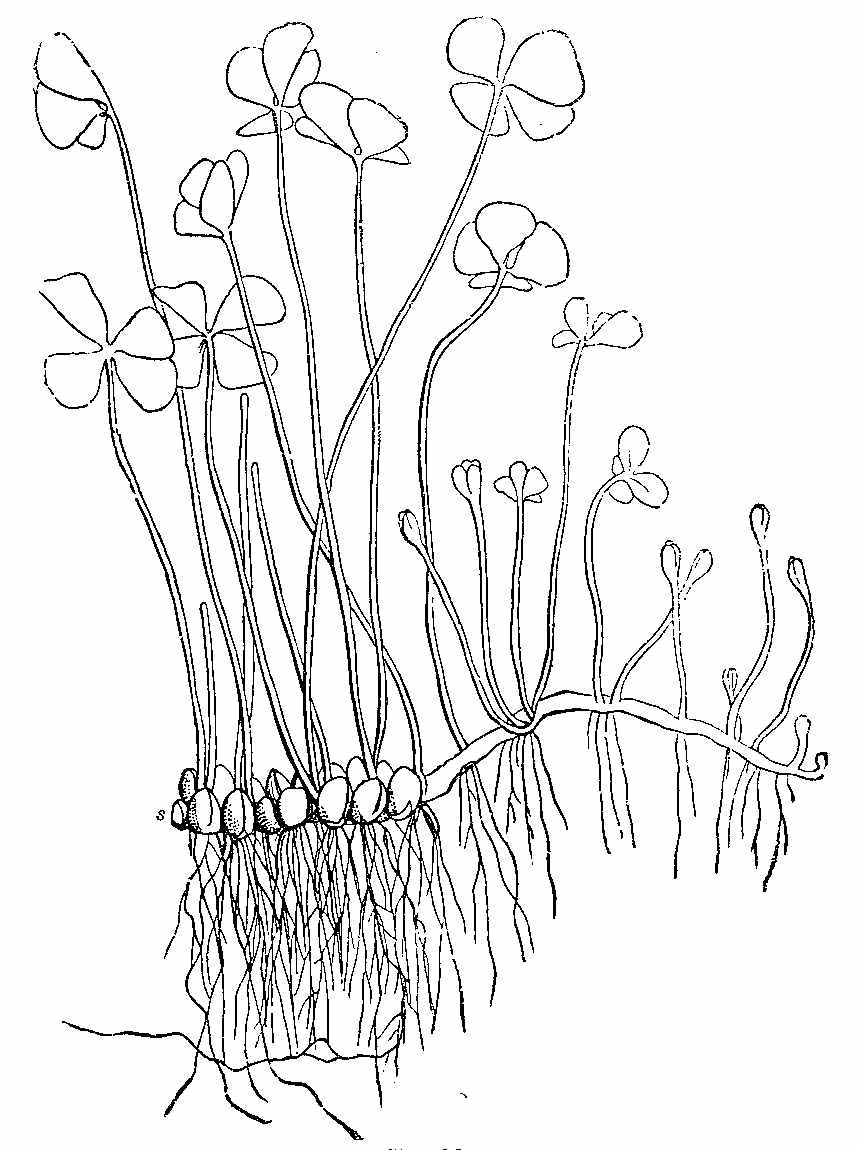
Among vascular Acrogens is included the natural order Marsileaceæ or Rhizocarpeæ, the Pepperworts (Fig. 22). The order consists of aquatic plants, with creeping stems, bearing leaves, which are either linear, or divided into three or more wedge-shaped portions not unlike clover. The fructification is at the base of the leaf-stalks, and consists of sacs (sporocarps) containing spores of two kinds, microspores and macrospores. The order contains Marsilea, Pilularia, Azolla, and Salvinia.
For a fuller account of Acrogenous plants, see Balfour's Class Book of Botany, p. 954.
These orders are represented in the Palæozoic flora. Many of the fossil species assume a large size, and show a greater degree of development than is seen in their recent congeners. The most important coal plants belong to the Ferns, Lycopods, and Horse-tails. The examination of the structure and conformation of the plants of the present flora assists much in the determination of the fossil carboniferous flora.
In the lower Palæozoic strata the plants which have been detected are few. In the Silurian and Cambrian systems, we meet with the remains of ancient marine plants, as well as a few terrestrial species. Even in the still older Laurentian rocks, if the remarkable structure known as Eozoon canadense[32] be considered, as it generally is, an animal, the existence of contemporary plants may be inferred, inasmuch as without vegetable life animals could not obtain food. In the Lower Silurian or Grauwacke, near Girvan, Hugh Miller found a species resembling Zostera in form and appearance. In the Lower Old Red Sandstone of Scotland he detected Fucoids, a Lepidodendron, and Lignite with a distinct Coniferous structure resembling that of Araucaria,[1] besides a remarkable pinnate frond. In the middle Old Red of Forfarshire, as seen in the Arbroath pavement, he found a fern with reniform pinnæ and a Lepidodendron. In the Upper Old Red, near Dunse, a Calamite and the well-known Irish fern Cyclopteris Hibernica occur.[2] This fern, Palæopteris [33]Hibernica of Schimper (Plate I. Figs. 1 to 4), along with Sigillaria dichotoma, is very abundant in beds of the same age in the south of Ireland, from which the specimens described by Edward Forbes were obtained. The fructification has recently been discovered. This shows that the fern belongs to[34] the Hymenophylleæ, and is consequently nearly related to the equally famous Killarney fern, Trichomanes radicans.
Mr. Carruthers states that the frond-stalk of this fern is thick, of considerable length, and clothed with large scales, which form a dense covering at the somewhat enlarged base. The well-defined separation observed in several specimens probably indicates that the frond-stalks were articulated to the stem or freely separated from it, and some root-like structures which occur on the slabs with the ferns may be their creeping rhizomes. The pinnæ are linear, obtuse, and almost sessile. The pinnules are numerous, overlapping, of an ovate or oblong-ovate form, somewhat cuneate below, and with a decurrent base. The veins are very numerous, uniform, repeatedly dichotomous, and run out to the margin, where they form a slight serration. Single pinnules rather larger than those of the pinnæ are placed over the free spaces of the rachis, as was pointed out by Brongniart. Carruthers has not met with any recent fern in which this occurs; but it has been observed in several fossil species, as in the allied American Palæopteris Halliana (Sch.), in Sphenopteris erosa (Morris), and others. The pinnules are sometimes entirely, but only partially fertile. The ovate-oblong sori are generally single and two-lipped, the slit passing one-third of the way down the sorus. The vein is continued as a free receptacle in the centre of the cup or cyst, as in existing Hymenophylleæ, in which it is included, not reaching beyond its entire portion. In some specimens the receptacle is broad or thick, indicating the presence of something besides itself in the cup, and giving the appearance that would be produced if it were covered with sporangia; there is no indication on the outer surface which might have been expected from the separate sporangia. The compression of the specimens in the rock, which has made the free receptacle appear like a vein on the wall of the cup, together with the highly altered condition of the rock in which the fossils are contained, accounts for the[35] imperfect preservation of the minute structures. The interpretation here given of the fructification of this interesting fossil exhibits so close a resemblance to what we find in the living genus Hymenophyllum, that, were it not for the vegetative portions, it would be placed in that genus. Several ferns have been described by Bunbury from Devonian rocks at Oporto. A still more extensive and varied land flora of Devonian age (or Erian, as he calls it) has been described and illustrated by Principal Dawson from the rocks of that period occurring in Canada; and during a recent visit to Britain he has correlated many of the fragments collected by Miller, Peach, and others, with the American species he has described. The following are some of the fossil plants from beds older than the Carboniferous system:[3]—Prototaxites Logani, Dadoxylon Ouangondianum, Calamites transitionis, Asterophyllites parvulus, Sphenophyllum antiquum, Lepidodendron Gaspianum, Lepidostrobus Richardsoni, L. Matthewi, Psilophyton princeps, P. robustius, Selaginites formosus, Cordaites Robbii, C. angustifolius, Cyclopteris Jacksoni.
From the microscopic examination of the structure of specimens of fossil trunks described under the name of Prototaxites Logani, and which Principal Dawson believes to be the oldest known instance of Coniferous wood, Mr. Carruthers has come to the conclusion that they are really the stems of huge Algæ, belonging to at least more than one genus. They are very gigantic when contrasted with the ordinary Algæ of our existing seas, nevertheless some approach to them in size is made in the huge and tree-like Lessonias which Dr. Hooker found in the Antarctic Seas, and which have stems about 20 feet high, with a diameter so great that they have been collected by mariners in these regions for fuel, under the belief that they were drift-wood. They are as thick as a man's [36]thigh. Schimper regards the Psilophyton of Dawson (Plate IV. Fig. 5) as allied to Pilularia, one of the Rhizocarps (Fig. 22), and Carruthers places it among the true Lycopodiaceæ.
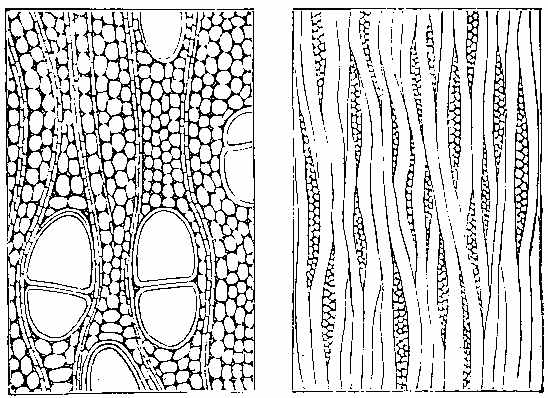
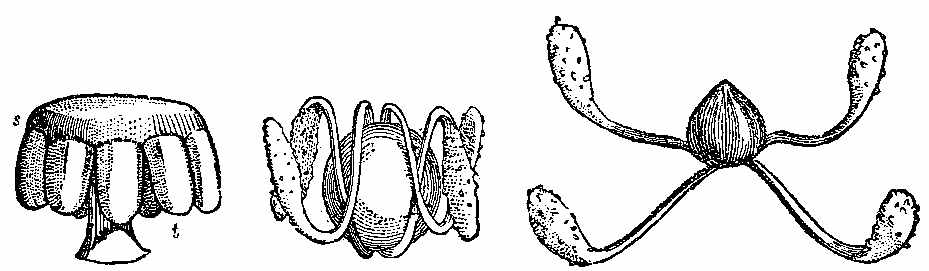
The Carboniferous period is one of the most important as regards fossil plants. The vegetable forms are numerous, and have a great similarity throughout the whole system, whether exhibited in the Old or the New World. The important substance called Coal owes its origin to the plants of this epoch. It has been subjected to great pressure and long-continued metamorphic action, and hence the appearance of the plants has been much altered. It is difficult to give a definition of Coal. The varieties of it are numerous. There is a gradual transition from Anthracite to Household and Parrot Coal; and the limit between Coal and what is called bituminous shale is by no means distinct. Coal may be said to be chemically-altered vegetable matter inter-stratified with the rocks, and capable of being used as fuel. On examining thin sections of coal under the microscope, we can detect vegetable tissues both of a cellular and vascular nature. In Wigan cannel coal, vegetable structure is seen throughout the whole mass. Such is likewise the case with other cannel, parrot, and gas coals. In common household coal, also, evident traces of organic tissue have been observed. In some kinds of coal punctated woody tissue (Plate III. Fig. 5) has been detected, in others scalariform tissue (Plate III. Fig. 6), as well as cells of different kinds. Sporangia are also frequently found in the substance of coal, as shown by Mr. Daw in that from Fordel (Plate III. Figs. 1 to 3); and some beds, like the Better bed of Bradford, are composed almost entirely of these sporangia imbedded in their shed microspores, as has been recently shown by Huxley. The structure of coal in different beds, and in different parts of the same bed, seems to vary according to the nature of the plants by which it has been formed, as well as to the metamorphic action which it has undergone. Hence the[37] different varieties of coal which are worked. The occurrence of punctated tissue indicates the presence of Coniferæ in the coal-bed, while scalariform vessels point to ferns, and their allies, such as Sigillaria and Lepidodendron. The anatomical structure of the stems of these plants may have some effect on the microscopic characters of the coal produced from them. In some cannel coals structure resembling that of Acrogens has been observed. A brownish-yellow substance is occasionally present, which seems to yield abundance of carburetted hydrogen gas when exposed to heat.
It appears that in general each bed of coal is accompanied by the remains of a somewhat limited amount of species. Their number, particularly in the most ancient beds, is scarcely more than eight or ten. In other cases the number is more considerable, but rarely more than thirty or forty. In the same coal-basin each layer often contains several characteristic species which are not met with either in the beds above or below. Thus, there are sometimes small local or temporary floras, each of which has given birth to layers of coal. The quantity of carbonaceous and other matter required to form a bed of coal is immense. Maclaren has calculated that one acre of coal three feet thick is equal to the produce of 1940 acres of forest.[4] The proportion of carbon varies in different kinds of coal. Along with it there is always more or less of earthy matter which constitutes the ashes. When the earthy substances are in such quantity that the coaly deposit will not burn as fuel, then we have what is called a shale. The coal contains plants similar to those of the shales and sandstones above and below it. Underneath a coal-seam lies the Underclay, containing roots only, and representing the ancient soil; then comes the Coal, composed of plants whose roots are in the clay, with others which have grown along with and upon them, in a manner precisely similar to the growth of peat at the present day; while above the coal is the Shale, [38]marking how mud was laid down on the plants, and bearing evidences of vigorous vegetation on neighbouring land, from which currents brought down the fine sediment, mingled with broken pieces of plants.
The total thickness of coal in the English coal-fields is about 50 or 60 feet. In the Mid-Lothian field there are 108 feet of coal. Coal-beds are worked at 1725 feet below the sea-level, and probably extend down to upwards of 20,000 feet. They rise to 12,000 feet above the sea-level, and at Huanuco, in Peru, to 14,700.[5] It is said that the first coal-works were opened at Belgium in 1198, and soon after in England and Scotland; it was not till the fifteenth century that they were opened in France and Germany.
The following calculations have been made as to the extent of the coal formation in different countries, and the amount of coal raised:—[6]
| Countries. | Square Miles of Coal Formation. | Annual Production of Coal in Tons. |
| Great Britain and Ireland | 5,400 | 65,887,900 |
| British North America | 7,530 | 1,500,000 |
| United States | 196,650 | 5,000,000 |
| Belgium | 518 | 8,409,330 |
| France | 1,719 | 7,740,317 |
| Prussia and Austria | —— | 4,200,000 |
| Saxony | 30 | 1,000,000 |
| Russia | 100 | 3,500,000 |
| Japan, China, Borneo, Australia, etc. | —— | 2,000,000 |
| Total Produce of the World | —— | 99,237,547 |
The total quantity of coal annually raised over the globe appears thus to be about 100 millions of tons, of which the produce of Great Britain is more than two-thirds, and would be sufficient to girdle the earth at the equator with a belt of 3 feet in thickness and nearly 5 feet in width. The coal-fields of the United States are nearly forty times larger than those of Great Britain.
Roscoe gives the following estimated quantities of coal in the principal countries:—
| Countries. | Average Thickness. No. Feet. | Tons. |
| Belgium | 60 | 36,000,000,000 |
| France | 60 | 59,000,000,000 |
| British Islands | 35 | 190,000,000,000 |
| Pennsylvania | 25 | 316,400,000,000 |
| Great Appalachian Coalfield | 25 | 1,387,500,000,000 |
| Indiana, Illinois, Western Kentucky | 25 | 1,277,500,000,000 |
| Missouri, and Arkansas Basin | 10 | 739,000,000,000 |
| North America (assumed thickness over an area of 200,000 square miles) | 20 | 4,000,000,000,000 |
Unger enumerates 683 plants of the coal-measures, while Brongniart notices 500. Of the last number there are 6 Thallogens, 346 Acrogens, 135 Gymnosperms, and 13 doubtful plants. This appears to be a very scanty vegetation, as far as regards the number of species. It is only equal to about 1/20th of the number of species now growing on the surface of the soil of Europe. Although, however, the number of species was small, yet it is probable that the individuals of a species were numerous. The proportion of Ferns was very large. There are between 200 and 300 enumerated. Schimper thinks there are 7 species congeneric with Lycopodium found in the coal-measures. The following are some of the Cryptogamous and Phanerogamous genera belonging to the flora of the Carboniferous period:—Cyclopteris, Neuropteris, Odontopteris, Sphenopteris, Hymenophyllites, Alethopteris, Pecopteris, Coniopteris, Cladophlebis, Senftenbergia, Lonchopteris, Glossopteris, Caulopteris, Lepidodendron (Lepidostrobus, Lepidophyllum, Knorria), Flemingites, Ulodendron, Halonia, Psaronius, Sigillaria and Stigmaria, Calamites (Asterophyllites and Sphenophyllum), Noeggerathia, Walchia, Peuce, Dadoxylon, Pissadendron, Trigonocarpum.
Ferns are the carboniferous fossil group which present the most obvious and recognisable relationship to plants of the present day. While cellular plants and those with lax tissues[40] have lost their characters by the maceration to which they were subjected before fossilisation took place, ferns are more durable, and retain their structure. It is rare, however, to find the stalk of the frond completely preserved down to its base. It is also rare to find fructification present. In this respect, fossil Ferns resemble Tree-ferns of the present day, the fronds of which rarely exhibit fructification. Hooker states that of two or three kinds of New Zealand Tree-fern, not one specimen in a thousand bears a single fertile frond, though all abound in barren ones. Only one surface of the fossil Fern-frond is exposed, and that generally the least important in a botanical point of view. Fructification is sometimes evidently seen, as figured by Corda in Senftenbergia. In this case the fructification is not unlike that of Aneimidictyon of the present day. Carruthers has recently detected the separate sporangia of Ferns full of spores in calcareous nodules in coal (Plate I. Fig. 5). These have the elastic ring characteristic of the Polypodiaceæ, and in their size, form, and method of attachment, they are allied to the group Hymenophylleæ. The absence of fructification presents a great obstacle to the determination of fossil Ferns. Circinate vernation, so common in modern Ferns, is rarely seen in the fossil species, and we do not in general meet with rhizomes. Characters taken from the venation and forms of the fronds are not always to be depended upon, if we are to judge from the Ferns of the present day. There is a great similarity between the carboniferous Ferns of Britain and America; and the same species, or closely allied species of the same genera as those found in Britain have been met with in South Africa, South America, and Australia. In the English coal-measures the species are about 140. The Palæozoic flora of the Arctic regions also resembles that of the other quarters of the globe. Heer, in his account of the fossil flora of Bear Island,[7] enumerates the following plants:—Cardiopteris frondosa, C. polymorpha,[41] Palæopteris Roemeriana, Sphenopteris Schimperi, Lepidodendron Veltheimianum, L. commutatum, L. Carneggiannum, L. Wilkianum, Lepidophyllum Roemeri, Knorria imbricata, K. acicularis, Calamites radiatus, Cyclostigma Kiltorkense, Stigmaria ficoides, etc., Cardiocarpum ursinum, C. punctulatum, besides various sporangia and spores.
The preponderance of Ferns over flowering plants is seen at the present day in many tropical islands, such as St. Helena and the Society group, as well as in extra-tropical islands, as New Zealand. In the latter, Hooker picked 36 kinds in an area of a few acres; they gave a luxuriant aspect to the vegetation, which presented scarcely twelve flowering plants and trees besides. An equal area in the neighbourhood of Sydney (in about the same latitude) would have yielded upwards of 100 flowering plants, and only two or three Ferns. This Acrogenous flora, then, seems to favour the idea of a humid as well as mild and equable climate at the period of the coal formation—the vegetation being that of islands in the midst of a vast ocean. Lesquereux, in Silliman's Journal, gives three sections of Ferns in the Carboniferous strata—viz. Neuropterideæ, Pecopterideæ, and Sphenopterideæ. In Neuropterideæ fructification has been seen in Odontopteris. In this genus the spores are in a peculiar bladdery sporangium. In Neuropterideæ the fructification appears to have resembled Danæa in some cases, and Osmunda in others. Professor Geikie has noticed in the lower Carboniferous shales of Slateford, near Edinburgh, a fern which has been named Adiantites Lindseæformis by Bunbury (Fig. 22, bis).[42] It has pinnules between crescent and fan shaped. (Mem. Geol. Survey of Edinburgh, 1861, p. 151.)
Among the Ferns found in the clays, ironstones, and sandstones of the Carboniferous period, we shall give the characters of some by way of illustration.[8] Pecopteris (Fig. 23) seems to be the fossil representative, if not congener, of Pteris. Pecopteris heterophylla (Fig. 24) has a marked resemblance to Pteris esculenta of New Zealand. The frond of Pecopteris is pinnatifid, or bi-tri-pinnatifid—the leaflets adhering to the rachis by the whole length of their base, sometimes confluent; the midrib of the leaflets runs to the point, and the veins come off from it nearly perpendicularly, and the fructification when present is at the end of the veins. Neuropteris (Figs. 25, 26, 27) has a pinnate or bipinnate frond, with pinnæ somewhat cordate at the base—the midrib of the pinnæ vanishing towards the apex, and the veins coming off obliquely, and in an arched manner. Neuropteris gigantea (Fig. 26) has a thick bare rachis, according to Miller, and seems to resemble much Osmunda regalis. Odontopteris has leaves like the last, but its leaflets adhere to the stalk by their whole base, the veins spring from the base of the leaflets, and pass on towards the point. Sphenopteris (Fig. 28) has a twice or thrice pinnatifid frond, the leaflets being narrowed at the base, often wedge-shaped, and the veins generally arranged as if they radiated from the base. Sphenopteris elegans resembled Pteris aquilina in having a stout leafless rachis, which divided at a height of seven or eight inches from its club-like base into two equal parts, each of which continued to undergo two or three successive bifurcations. A little below the first forking two divided pinnæ were sent off. A very complete specimen, with the stipe, was collected in the coalfield near[43] Edinburgh by Hugh Miller, who has described it as above. Lonchopteris has its frond multi-pinnatifid, and the leaflets more or less united together at the base; there is a distinct midrib, and the veins are reticulated. Cyclopteris (Fig. 29) has simple orbicular leaflets, undivided or lobed at the margin, the veins radiating from the base, with no midrib. Schizopteris resembles the last, but the frond is deeply divided into numerous unequal segments, which are usually lobed and taper-pointed.
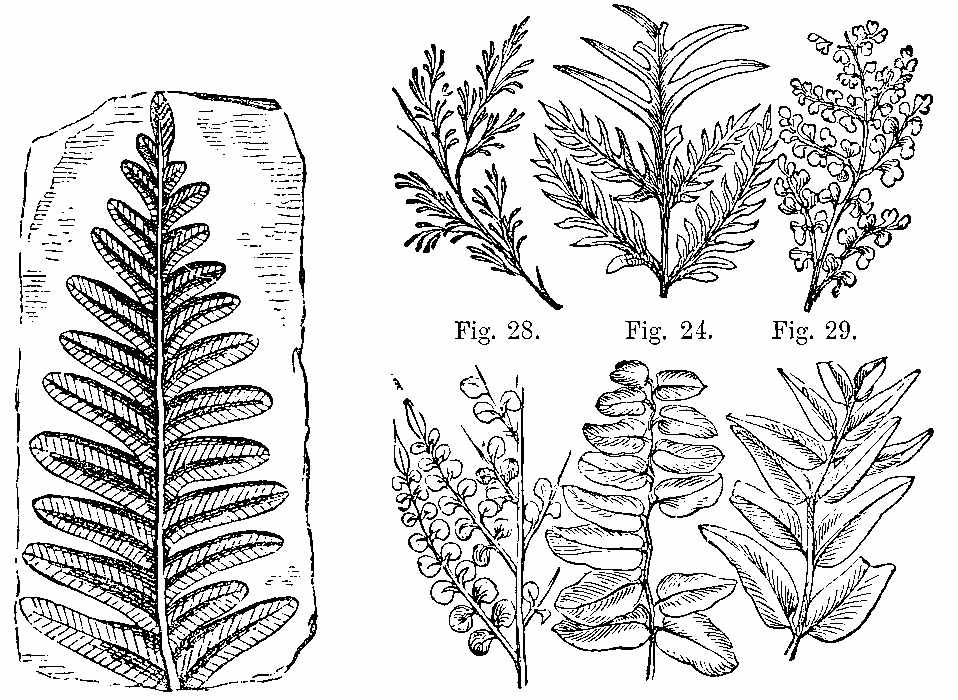

The rarity of Tree-ferns in the coal-measures has often been observed, and it is the more remarkable from the durable nature of their tissues. Several species have, however, been noticed. They are referred to the genus Caulopteris. One of them, C. macrodiscus (Fig. 30) has the leaf-scars in linear series. Two other species are figured, the one a slender form[44] with the scars widely separated, as in some Alsophilas, C. Balfouri (Fig. 31) from the Somersetshire coal-field; and the other with larger stems and more closely aggregated scars, C. Morrisi (Fig. 32), from the coal-measures at Newcastle. The latter species shows the cavities at the base of the petiole described by Mohl in many living fern-stems. The fossils named Psaronius appear to have been fern-stems with a slender axis and a large mass of adventitious roots, as in some Dicksonias and in Osmunda regalis. These stems probably belong to some of the fronds to which other names are given, but as they have not been found attached, it is impossible to determine the point. Miller has described a fern as occurring in the coal-measures, which at first sight presents more the appearance of a Cycadaceous frond than any other vegetable organism of the carboniferous age except the Cycadites Caledonicus (Salter), from Cockburnspath Cove. He thus describes it:—
"From a stipe about a line in thickness there proceed at[45] right angles, and in alternate order, a series of sessile lanceolate leaflets, rather more than two inches in length, by about an eighth part of an inch in breadth, and about three lines apart. Each is furnished with a slender midrib; and, what seems a singular, though not entirely unique feature in a Fern, the edges of each are densely hirsute, and bristle with thick short hair. The venation is not distinctly preserved."
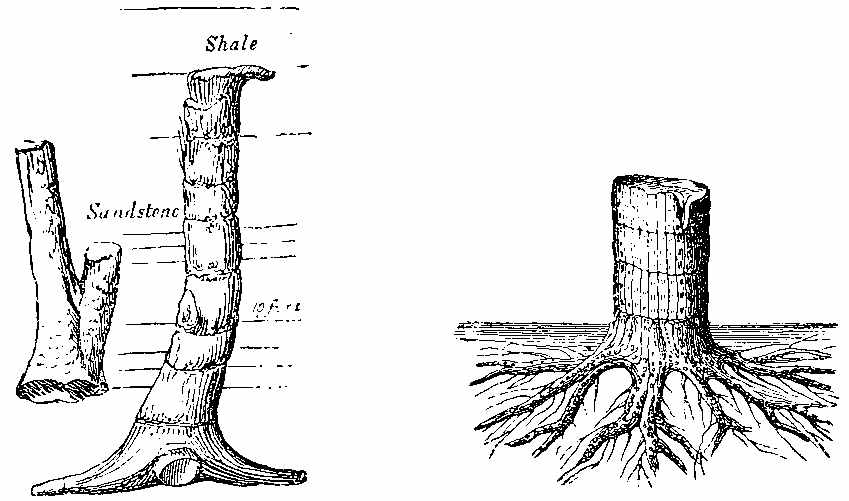
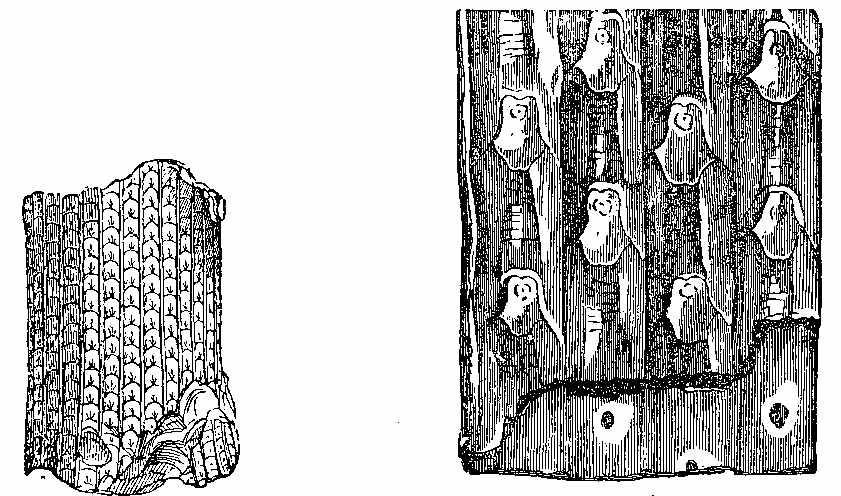
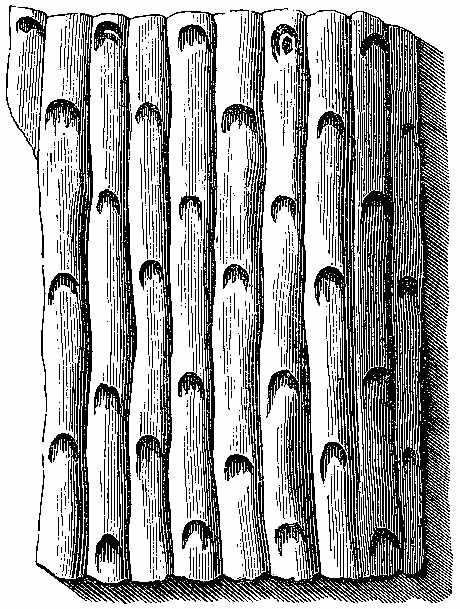
Sigillaria (Plate IV. Figs. 1 and 2) is perhaps the most important plant in the coal formation. The name is derived from sigillum, a seal, to indicate the seal-like markings in the stem. It is found in all coal-shales over the world. Schimper mentions 83 species. It occurs in the form of lofty stems, 40-50 feet high, and 5 feet broad (Figs. 33 and 34). Many stems of Sigillaria may be seen near Morpeth, standing erect at right angles to the planes of alternating strata of shale and sandstone (Fig. 33). They vary from 10 to 20 feet in height, and from one to three feet in diameter. Sir W. C.[46] Trevelyan counted 20 portions of these trees within the length of half-a-mile, of which all but four or five were upright. Brongniart mentions similar erect stems as being found near St. Etienne. The stem of Sigillaria is fluted in a longitudinal manner, like a Doric column, and has a succession of single scars, which indicate the points of insertion of the leaves (Figs. 35, 36, and 37). When the outer part of the stem separates like bark, it is found that the markings presented by the inner surface differ from those seen externally. This has sometimes given rise to the erroneous multiplication of species and even of genera.[47] Sigillaria elegans, as figured by Brongniart in Archives du Museum, i. 405, has a stem consisting of a central cellular axis or medulla, surrounded by a vascular cylinder, and this is invested by a thick cellular cortical layer, the outer portion composed of fusiform cells of less diameter than those of the inner portion. What Brongniart calls medullary rays are mere cracks or separations in the wedges traversed by vessels. In its structure it resembles its root Stigmaria, and must be referred to Lycopodiaceæ, along with Lepidodendron, Halonia, Ulodendron, etc. The small round sporangia of Sigillaria are borne in a single patch on the somewhat enlarged bases of some of the leaves. (See Carruthers on Structure and Affinities of Sigillaria, in Journ. Geol. Soc. Aug. 1869.)
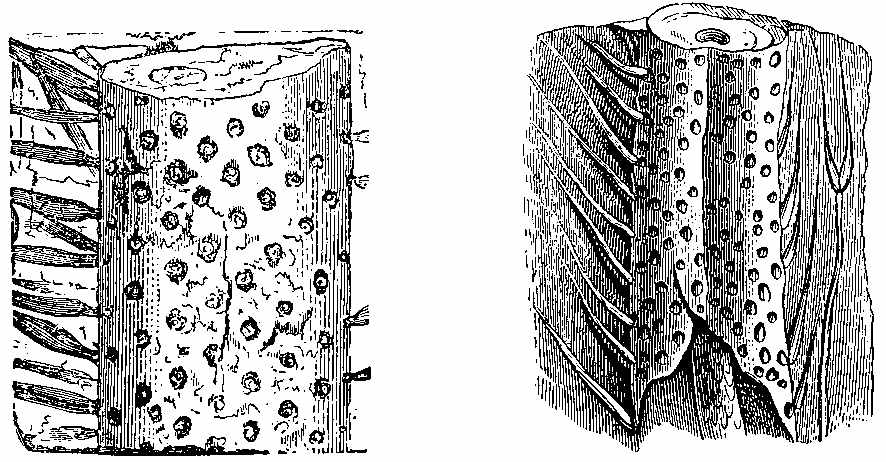
It has been ascertained by Professor King and Mr. Binney of Manchester, that the plant called Stigmaria (Fig. 38) is not a separate genus, but the root of Sigillaria (Plate IV. Figs. 1 and 2). The name is derived from στίγμα, a mark, indicating the markings on the axis. It is one of the most common productions of the coal-measures, and consists of long rounded or compressed fragments, marked externally by shallow circular,[48] oblong, or lanceolate cavities (Fig. 39) in the centre of slight tubercles, arranged more or less regularly in a quincuncial manner (Plate III. Fig. 7). The cavities occasionally present a radiating appearance. The axis of the fragments is often hollow, and different in texture from the parts around. This axis consists of a vascular cylinder or woody system, penetrated by quincuncially arranged meshes or openings, through which the vascular bundles proceed from the inner surface of the cylinder to the rootlets (Plate III. Figs. 8 and 9). From the scars and tubercles arise long ribbon-shaped processes, which were cylindrical cellular roots, now compressed (Fig. 38). The vascular cylinder of Stigmaria is composed entirely of scalariform tissue, pierced by meshes for the passage, from the inner surface of the cylinder, of the vascular bundles which supply the rootlets. (Carruthers in Geol. Proc., Aug. 1869.) Stigmaria ficoides (Fig. 38) abounds in the under-clay of a coal-seam, sending out numerous roots from its tubercles, and pushing up its aerial stem, in the form of a fluted Sigillaria. On the Bolton and Manchester Railway Mr. Binney discovered Sigillarias standing erect, and evidently connected with Stigmarias which extended 20 feet or more.[9] Stigmaria is regarded by Schimper as roots, not of Sigillaria only, but of Knorria longifolia (one of the Lepidodendreæ). The base of the stem of this species of Knorria is Ancestrophyllum, and the upper part is Didymophyllum Schottini of Goeppert. Professor King and others suppose that the Fern-like frond called Neuropteris is connected with Sigillaria, but this is a mere conjecture, set aside by the discovery of leaves attached to a species allied to Sigillaria elegans, which establishes that the long linear leaves described under the name Cyperites are the foliage of this genus. Goldenberg has figured the fructification, which consists of small sporangia like those of Flemingites,[49] borne on the basis of but slightly modified leaves. This establishes the opinion that Sigillaria was an acrogenous plant belonging to Lycopodiaceæ. Brongniart reckons it as representing an extinct form of Gymnosperms, and King, having erroneously associated the Cyclopteris with it, places it between the Ferns and Cycadaceæ. Mr. Carruthers informs me that he has examined the stem of a true fluted Sigillaria, with the tissues preserved, and that these agree with the structure of Lepidodendron, a position in which he had already placed it from the structure of its fruit.
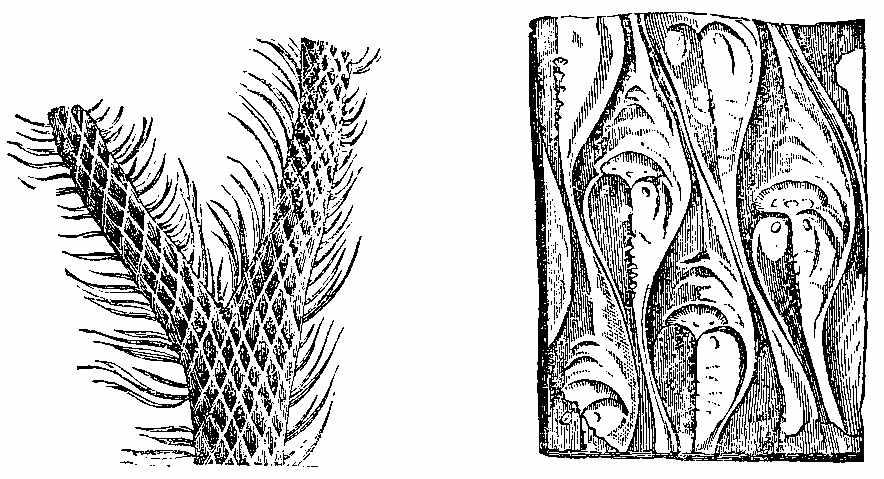


Lepidodendron (Figs. 40 to 44) is another genus of the coal-measures which differs from those of the present day (Plate IV. Fig. 3). Lepidodendrons, or fossil Lycopodiaceæ, had spikes of fructification comparable in size to the cones of firs and cedars, and containing very large sporangia, even larger than those of Isoetes, to which they approach in form and structure. Schimper, in 1870, enumerates 56 species of Lepidodendron, all arborescent and carboniferous. The stem of a Lepidodendron is from 20 to 45 feet high, marked outside[50] by peculiar scale-like scars (Fig. 41), hence the name of the plant (λεπίς, a scale, and δένδρον, a tree). Although the scars on Lepidodendron are usually flattened, yet in some species they occupy the faces of diamond-shaped projections, elevated one-sixth of an inch or more above the surface of the stem, and separated from each other by deep furrows;—the surface bearing the leaf being perforated by a tubular cavity, through which the bundle of vessels that diverged from the vascular axis of the stem to the leaf passed out. The linear or lanceolate leaves are arranged in the same way as those of Lycopodiums or of Coniferæ, and the branches fork like the former. The internal structure of the stem is the same as that of Sigillaria. The fruit of Lepidodendron and allied genera is seen in Lepidostrobus and Triplosporites (Figs. 42, 43; Plate III, Fig. 10). [51]Carruthers, in his lecture to the Royal Institution, in describing the forms of Lepidostrobus, says—"The fruit is a[52] cone composed of imbricated scales arranged spirally on the axis like the true leaves, and bearing the sporangia on their horizontal pedicels. Three different forms of fruit belong to this genus, or it should perhaps rather be called group of plants. The first of these is the cone named by Robert Brown Triplosporites (Figs. 42, 43), and described by him from an exquisitely preserved specimen of an upper portion, in which the parts are exhibited as clearly in the petrified condition as if they belonged to a fresh and living plant. The large sporangia have a double wall, the outer composed of a compact layer of oblong cells placed endwise, or with the long diameter perpendicular to the surface; the inner is a delicate cellular membrane. The sporangium is filled with a great number of very small spores, each composed of three roundish bodies or sporules. Recently Brongniart and Schimper have described a complete specimen of this fruit, in which the minute triple spores are confined to the sporangia of the upper and middle part of the cone, but the lower portion, which was wanting in Brown's specimen, bears sporangia filled with simple spherical spores ten or twelve times larger than the others (woodcut 44, 9).
"The structure of another form of cone (Lepidostrobus) has been expounded by Dr. Hooker. The arrangement of the different parts comprising it is precisely similar to what occurs in Triplosporites; but the sporangia are filled with the minute triple spores throughout the whole cone (woodcut 44, 6 and 8).
"The third form of cone, described by me under the name Flemingites, differs from the other two in having a large number of small sporangia supported on the surface of each[53] scale; and it agrees with Lepidostrobus in the sporangia containing only small spores (woodcut 44, 10).
"In comparing these fossils with the living club-mosses, one is struck with the singular agreement in the organisation of plants so far removed in time, and so different in size, as the recent humble club-mosses and the palæozoic tree Lepidodendrons. The fruit of Triplosporites, like that of Selaginella (woodcut 44, 1), contains large and small spores, the microspores being found in both genera on the middle and upper scales of the cone, and the macrospores on those of the lower portion (Fig. 43).
"On the other hand, the fruits of Lepidostrobus and Flemingites agree with that of Lycopodium in having only microspores. The size of the two kinds of spores also singularly agrees in the two groups. This is of some importance, for among the recent vascular Cryptogams there is a remarkable uniformity in the size of the spores in the members of the different groups, even when there is a great variety in the size of the plants. Thus the spore of our humble wall-rue is as large as that of the giant Alsophila of tropical regions. So also the spores of Equisetum and Calamites agree in size, as may be seen in woodcut 47, Figs. 3, 4, and 9, where the spores of the two genera are magnified to the same extent. And a similar comparison of the macrospore and microspore of Triplosporites with those of Selaginella, and of the microspore of Lepidostrobus with that of Lycopodium, exhibits a similar agreement. This is made apparent by the drawings in woodcut 44 of the two kinds of spores of Selaginella, 3 and 4, with those of Triplosporites, 8 and 9, which are drawn to the same scale."
The genus Sigillaria, as we have already said, has, according to the observation of Hooker, small sporangia exactly agreeing in size and form with those of Flemingites. Most probably the contents of these small sporangia were the same in both genera, so that Sigillaria would be placed with Flemingites[54] and Lepidostrobus as arborescent Lycopodiaceæ having their affinities with Lycopodium, as they have all microspores only in their fructification.
The scales upon the Lepidodendron stems, as well as those in the cones, are arranged in a spiral manner, in the same way as plants of the present day. Professor Alexander Dickson has examined the phyllotaxis of Lepidodendrons, and gives the following results of his observations (Trans. Bot. Soc. Edin. xi. 145). The fossil remains of Lepidodendrons are often so compressed that it is difficult, or even impossible, to trace the secondary spirals round the circumference of the stem. In those cases, however, where there is comparatively little compression, i.e. where the stem is more or less cylindrical, the determination of the phyllotaxis is easy. Of such stems he has examined fifteen specimens, which may be classed according to the series of spirals to which the leaf-arrangement belongs:—
A. Ordinary series, ½, ⅓, 2/5, ⅜, 5/13, etc.
(a.) Single spirals (D turning to the right, S to the left).
(1.) Lepidodendron (Possil Ironstone series). Stem about ¾ of an inch in diameter. Secondary spirals 8 D, 13 S, 21 D. Divergence = 13/34 (or possibly 21/55).
(2.) Lepidodendron (Knightswood, near Glasgow, Mr. J. Young). Stem about 1½ inch in diameter. Secondary spirals 13 D, 21 S, 34 D. Divergence = 21/55.
(3.) Lepidodendron (Possil Sandstone series). Trunk about 2 feet long, with an average diameter of 20 inches. Steepest secondary spirals 55 S, 89 D. Divergence = 55/144.
(b.) Conjugate spirals.[11]
(4.) Lepidostrobus ornatus (Bathgate coal-field). About ¾ of an inch in diameter. Secondary spirals 10 D, 16 S, 26 D, 42 S. Divergence = 13/(34×2) (Bijugate arrangement).
[55](5.) Lepidostrobus (Plean, Stirlingshire, Mr. Mackenzie). About ½ an inch in diameter. Secondary spirals 9 S, 15 D, 24 S, 39 D. Divergence = 8/(21×3) (Trijugate arrangement).
(6.) Knorria taxina (from collection of Dr. Rankin, Carluke). Somewhat compressed, 2-2½ inches[12] in diameter. Secondary spirals 15 D, 24 S. Divergence = 15/(13×3) (Trijugate arrangement).
(7.) Lepidodendron (from Dr. Rankin's collection). About 1¼ inch in diameter. Secondary spirals 10 D, 15 S, 25 D, 40 S. Divergence = 5/(13×5) (Quinquejugate arrangement).
(8.) Lepidodendron (Dowanhill, Glasgow, Possil Sandstone series). Trunk about 1 foot long, and 1 foot in diameter. The upper portion exhibits secondary spirals 35 D, 56 S, 91 D; thus indicating a 7-jugate arrangement, with divergence = 8/(21×7). The arrangement on the middle and lower portion is indistinct and confused; so much so as to render any determination of the arrangement doubtful.
B. Series, ⅓, ¼, 2/7, 3/11, etc.
(9.) Lepidodendron (Messrs Merry and Cunningham's Clayband Iron-Pit, Carluke). Stem 2 inches in diameter. Secondary spirals 18 S, 29 D, 47 S. Divergence = 21/76.
C. Series, ¼, 1/5, 2/9, 3/14, etc.
(10.) Lepidodendron (R. B. Garden, Edinburgh, Museum). Stem somewhat flattened, 1-1½ inch in diameter. Secondary spirals 9 D, 14 S, 23 D, 37 S. Divergence = 13/60.
(11.) Lepidodendron (Redhaugh, near Edinburgh, Mr. Peach). Stem somewhat flattened, ¾ to ½ inch in diameter. Secondary spirals 9 S, 14 D, 23 S, 37 D. Divergence = 13/60.
D. Series, 1/5, 1/6, 2/11, 3/17, 5/28, etc.
(12.) Knorria taxina (Stockbriggs, Lesmahagow,—Hunterian Museum). About 1 inch in diameter. The specimen consists of a main stem and one of the branches into which it has forked. On the main stem the secondary spirals are 6 D, 11 S, 17 D. Divergence = 5/28 (series, 1/5, 1/6, 2/11, 3/17, 5/28, etc.)—On the branch the secondary spirals are 8 S, 13 D. Divergence = 8/21 (ordinary series, ½, ⅓, 2/5, ⅜, etc.)
E. Series, ½, 2/5, 3/7, 5/12, 8/19, 13/31, 21/50, etc.
(13.) Lepidodendron (from Dr. Rankin's collection). About ⅞ inch in diameter. Secondary spirals 12 D, 19 S, 31 D. Divergence = 21/50.
F. Series, ⅓, 3/10, 4/13, 7/23, 11/36, 18/59, etc.
(14.) Lepidodendron elegans (Possil Ironstone). About 1¼ inch in diameter. Secondary spirals 10 S, 13 D, 23 S, 36 D. Divergence = 18/59.
(15.) Lepidodendron (Possil Ironstone). About 2¼ inches in diameter. Secondary spirals 23 S, 36 D, 59 S, 95 D. Divergence = 47/154.
From the above it is evident that the phyllotaxis of Lepidodendron is extremely variable, as much so perhaps as that of those most variable plants, in this respect, the Cacti. It is also clear that what has been enunciated by Professor Haughton (Manual of Geology, Lond. 1866, pp. 243, 245) as the law according to which the leaves of palæozoic plants were arranged—viz. that of alternate whorls—does not apply to these ancient Lycopods. Lepidodendron aculeatum is noted by Naumann as exhibiting an 8/21 arrangement. (Poggendorff, Annalen, 1842, p. 5.) Professor Alexander Braun (Nov. Acta Ac. C. L. C. xv. 1, pp. 558-9), speaking of the excessive deviation from ordinary arrangements in Equisetaceæ (including Calamites), compares them in this respect with Lycopodiaceæ (including Lepidodendron), saying that in these two families "the utmost limits of the domain of all leaf-arrangement appears to be attained."
Lepidophyllum is certainly leaves of Lepidodendron, the different Lepidophylla belonging to different species of the genus. The slender terminal branches are noticed under the name of Lycopodites. In coal from Fordel Mr. Daw has detected innumerable bodies (Plate III. Figs. 1, 2, 3) which have been shown to be sporangia. (Balfour, Trans. Roy. Soc. Ed. xxi. 187.) On their under surface Mr. Carruthers has observed a triradiate ridge (Plate III. Fig. 4). (Geological Magazine, 1865, vol. ii. p. 140.) These sporangia have been[57] found connected with the cone-like fructification called Flemingites, and resembling Lycopodium (woodcut 44, Fig. 4). Many forms of fossil plants, such as Halonia, Lepidophloios, Knorria, and Ulodendron, belong to the Lepidodendron group. Knorria is said to be the internal cast of a Lepidodendron.
Ulodendron minus and U. Taylori (Plate III. Fig. 11), found in ferruginous shale in the Water of Leith, near Colinton, exhibit beautiful sculptured scars, ranged rectilinearly along the stem. The surface is covered with small, sharply relieved obovate scales, most of them furnished with an apparent midrib, and with their edges slightly turned up. The circular or oval scars of this genus are probably impressions made by a rectilinear range of aerial roots placed on either side. When decorticated, the stem is mottled over with minute dottings arranged in a quincuncial manner, and its oval scars are devoid of the ordinary sculpturings. Bothrodendron is a decorticated condition of Ulodendron.


Calamites (κάλαμος, a reed) is a reed-like fossil, having a sub-cylindrical jointed stem (Fig. 45, a and b; Fig. 46; Plate IV. Fig. 4). The stem is often crushed and flattened, and was originally hollow. Calamites is thus defined by Grand d'Eury (Ann. Nat. Hist. ser. 4, vol. iv. p. 124):—Stem articulated, fistular, and septate; outer part comparatively thin, formed of three concentric zones—1, an exterior cortical layer now converted into coal; 2, a thin subjacent zone of vascular tissue, now invariably destroyed; 3, a sort of inner lining epidermis, which is carbonified. Cortical envelope marked interiorly with regular flutings, interrupted and alternate at the articulations. Inner epidermis smooth,[58] or scarcely striated. Vascular cylinder thin; outer surface of bark more fully fluted and articulated than the inner surface.

Carruthers gives the following description of the structure of a species of Calamite which he examined:—The stem was composed of a central medulla, which disappeared with the growth of the plant, surrounded by a woody cylinder, composed entirely of scalariform vessels, and a thin cortical layer. The medulla penetrated the woody cylinder by a series of regular wedges, which were continued, as delicate laminæ of one or two cells in thickness, to the cortical layer. The cells of those laminæ were not muriform; their longest diameter was in the direction of the axis. The wedges were continuous, and parallel between each node. As the axial appendages were produced in whorls, the only interference with the regularity of the tissues was by the passing out through the stem at the nodes of the vascular bundles which supplied these appendages. As the leaves of each whorl were (with one or[59] two exceptions) opposite to the interspaces of the whorls above and below, there was also at each node a re-arrangement of the wedges of vascular and cellular tissues.
Schimper considers Calamites as having an analogy with Equisetum in its fructification. He looks on them as fossil Equisetaceæ. Annularia and Sphenophyllum are considered as establishing a passage from the Equisetaceæ to the Lycopodiaceæ. Some gigantic fossil Equiseta had a diameter of nearly 5 inches, and a height of 30 or more feet. The branches, which adorned the higher part of them in the form of a crown, are simple, and have at their extremity a spike of the size of a pigeon's egg, and organised exactly like the spikes of living Equiseta. The subterranean rhizomes are well developed, and gave origin, like many Equiseta, to tubercles which had the form and size of a hen's egg.
The characters of Equisetum of the present day and Calamites, are exhibited in woodcut 47. They show a marked resemblance in the fructification. (See also page 31.)
Plants of Calamites have been seen erect by Mr. Binney, and he has determined that what were called leaves or branches by some are in reality roots. Mr. Binney gives a full description of various Calamites, under the name of Calamodendron commune, in his Memoir published by the Palæontographical Society, 1868. There are between 50 and 60 species recorded.[13]
In Spitzbergen, in rocks of the Carboniferous epoch, there have been found Calamites, Sigillaria, Lepidodendron, and ferns, apparently the same as those found in the Carboniferous epoch in Europe—Calamites radiatus, Lepidodendron Veltheimianum, Sigillaria distans, Stigmaria ficoides. Some species—Sigillaria Malmgreni, Lepidodendron Carneggiannum, and L. Wilkianum—seem to be peculiar to Bear Island.
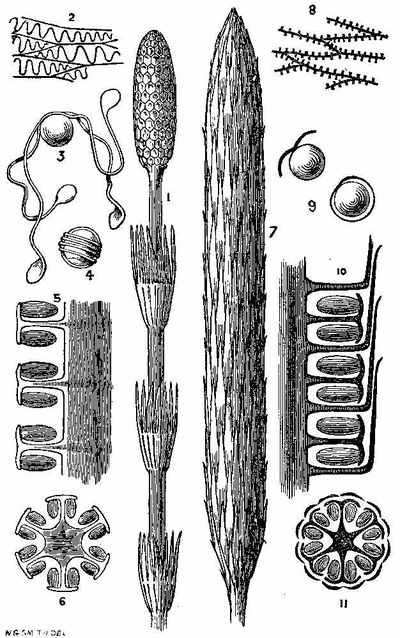
According to Carruthers the Equisetaceæ are represented in Britain by the two genera Calamites found in primary beds,[60] and Equisetum found in secondary rocks and living at the present day. The difference in the structure of their fruits is shown in woodcut 47. The fruit of Calamites, called Volkmannia Binneyi (woodcut 47, 7), is a small slender cone composed of alternating whorls of imbricate scales, twelve in each verticil. The scales completely conceal[61] the leaves connected with the fructification. The fruit-bearing leaves are stalked, peltate, and are arranged in whorls of 6. There are four sporangia borne on the under-surface of the peltate leaves. These spore-cases have cellular parietes, and in their interior there is a deposit of cellulose in the form of short truncate processes not unlike imperfect spirals. The spores are spherical, and appear to have thread-like processes proceeding from them, similar to elaters. The fruit-cone bears a marked resemblance to the fruit of Equisetum in its fruit-bearing leaves, sporangia, spores, and elaters (see Figs. 18, 19, 20, 21). In the modern plant all the leaves of the cone are fructiferous, while in the fossil plant some are fruit-bearing, and others are like the ordinary leaves of the plant. It is thought that the fossil may be reckoned as having a somewhat higher position than that possessed by the living genus.

The different forms of foliage called Asterophyllites, Sphenophyllum, and Annularia, belong to the one genus Calamites, but they may form, perhaps, well-characterised sections when their fruits are better known. In woodcut 48 representations are given of the foliage and fruit of varieties of Calamites. In 1 and 2 we see the simplest form called Asterophyllites. The leaves are linear and slender, with a single rib. The form called Annularia (3 and 4) differs chiefly in having a larger amount of cellular tissue spread out on either side of the midrib. This form has a different aspect in a fossil state from the other, from its whorls of numerous broad leaves spread out on the surface of deposition, while the acicular leaves of Asterophyllites have penetrated the soft mud, and are generally preserved in the position they originally occupied in reference to the supporting branch. The third form (5 and 6) is called Sphenophyllum, and consists of whorls of wedge-shaped leaves, with one or more bifurcating veins. They occur like those of Annularia, spread out on the surface of the shale.
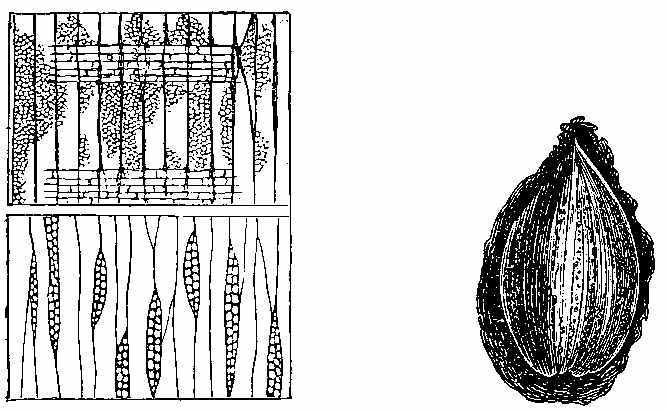
True Exogenous trees exist in the coal-fields both of England and Scotland, as at Lennel Braes and Allan Bank, in Berwickshire; High-Heworth, Fellon, Gateshead, and Wideopen, near Newcastle-upon-Tyne; and in quarries to the west of Durham; also in Craigleith quarry, near Edinburgh, and in the quarry at Granton, now under water. In the latter localities they lay diagonally athwart the sandstone strata, at an angle of about 30°, with the thicker and heavier part of their trunks below, like snags in the Mississippi. From their direction we infer that they have been drifted by a stream which has flowed[63] from nearly north-east to south-west. At Granton, one of the specimens exhibited roots. In other places the specimens are portions of stems, one of them 6 feet in diameter by 61 feet in length, and another 4 feet in diameter by 70 feet in length. These Exogenous trees are Gymnosperms, having woody tissue like that of Coniferæ. We see under the microscope punctated woody tissue, the rows of disks being usually two, three, or more, and alternating. They seem to be allied in these respects to Araucaria and Eutassa (Fig. 61, p. 74) of the present flora. Araucarioxylon or Pinites Withami (Fig. 49) is one of the species found in Craigleith quarry; the concentric layers of the wood are obsolete; there are 2, 3, or 4 rows of disks on the wood, and 2-4 rows of small cells in the medullary rays. Along with it there have also been found Dadoxylon medullare, with inconspicuous zones, 2, 3, and 4 rows of disks, and 2-5 series of rows of cells in the rays. Pissadendron antiquum (Pitus antiqua) having[64] 4-5 series of cells in the medullary rays, and P. primævum (Pitus primæva), with 10-15 series of cells in the medullary rays, occur at Tweedmill and Lennel Braes in Berwickshire; Peuce Withami (Fig. 1, p. 3) at Hilltop, near Durham, and at Craigleith. Sternbergia is considered by Williamson as a Dadoxylon, with a discoid pith like that seen now-a-days in the Walnut, Jasmine, and Cecropia peltata, as well as in some species of Euphorbia.[14] Sternbergia approximata is named by him Dadoxylon approximatum. Hooker believes from the structure of Trigonocarpum (Fig. 50) that it is a coniferous fruit nearly allied to Salisburia (Trans. Roy. Soc. 1854). Several species of Trigonocarpum occur in the Carboniferous rocks, such as T. olivæforme from Bolton (Plate II. Fig. 5), and T. sulcatum from Wardie, near Edinburgh (Plate II. Fig. 6). Noeggerathia and a few other plants, such as Flabellaria and Artisia, are referred by Brongniart to Cycadaceæ. Flabellaria borassifolia, according to Peach, has leaves like Yucca. Noeggerathia has pinnate leaves, cuneiform leaflets, sometimes fan-shaped; the veins arise from the base of the leaflets, are equal in size, and either remain simple or bifurcate, the nervation (venation) being similar to that of some Zamias.
The fossils of this period, referred to as Antholithes,[15] have just been shown by Mr. Carruthers to be the inflorescence of Cardiocarpum (Geol. Mag. Feb. 1872), and he proposes to set aside the former name, confining it to the tertiary fossils to which it was originally given by Brongniart, and to use the latter name. The main axis of the inflorescence is simple, stout, and marked externally with interrupted ridges. The [65]axis bears in a distichous manner sub-opposite or alternate bracts of a linear-lanceolate form and with decurrent bases. In the axils of the bracts were developed flower-like leaf-bearing buds, and from them proceeded three or four linear pedicels, which terminated upwards in a somewhat enlarged trumpet-shaped apex. To this enlarged articulating surface was attached the fruit, to which has been given the generic name Cardiocarpum[16] (Fig. 51). The place of attachment is indicated by the short straight line which separates the cordate lobes at the base of the fruit. The fruit is flattish, broadly ovate, with a cordate base and sub-acute apex. It consists of an outer pericarp, inclosing an ovate-acute seed. That the pericarp was of some thickness, and formed probably a sub-indurated rind, is shown by a specimen preserved in the round, and figured (Fig. 53 a). The pericarp is open at the apex; and the elongated tubular apex of the spermoderm passes up to this opening. The seed forms a distinct swelling in the centre of the fruit, and a slight ridge passes up the middle to the base of the apical opening.

These fossils are believed to be an extinct form of Gymnosperms. Two species have been described, of both of which[66] we are able to give figures. The first figure is from the specimens collected by Mr. Peach at Falkirk. It is Cardiocarpum Lindleyi (Figs. 51, 52); it has a primary axis with sub-opposite axillary axes, bearing four to six lanceolate leaves and three or four pedicels. Primary bracts short and arcuate. Fruit ovate-cordate, with an acute bifid apex, and a ridge passing up the middle of the fruit.

The second species is Cardiocarpum anomalum (Fig. 53) from Coalbrookdale; it has a primary axis with alternate or sub-opposite axillary axes, slender and elongated, bearing many linear leaves, and several slender pedicels; primary bracts long, slender, and straight; fruits small, margined. The somewhat magnified separate fruit (a) shows the thickness of the pericarp and the enclosed seed.
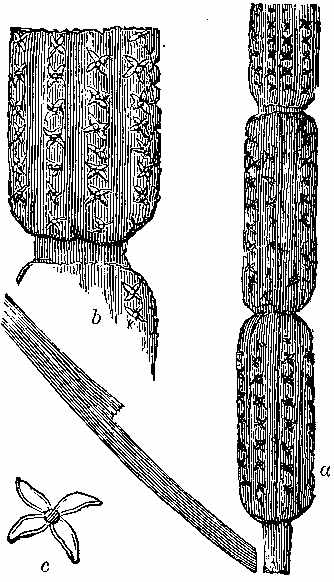
In the bituminous shale at Granton, near Edinburgh, Dr. Robert Paterson discovered in 1840 a peculiar fossil plant, which he called Pothocites Grantoni (Fig. 54, a). It is figured in the Transactions of the Edinburgh Botanical Society, vol. i. March 1840. It is a spike covered by parallel rows of flowers (Fig. 54, b), each apparently with a 4-cleft calyx (Fig. 54, c). It was supposed to be allied to Potamogeton or Pothos, more probably to the latter. In that case it must be referred to the natural order Araceæ. The original specimen is deposited in the museum at the Royal Botanic Garden, Edinburgh.
Our knowledge of the real state of the vegetation of the[67] earth when coal was formed must be very limited, when we reflect how seldom the fructification of coniferous trees has been met with in the coal-measures. A very doubtful fragment, supposed to be a cone, is given in Lindley and Hutton's work, under the title of Pinus anthracina; but it is believed by Carruthers to be a fragment of a Lepidodendroid branch. Lyell never saw a fossil fir-cone of the Carboniferous epoch, either in the rocks or museums of North America or Europe. Bunbury never heard of any other example than that noticed by Lindley and Hutton. Principal Dawson is disposed to think that the suberin of cork, of epidermis in general, and of spore-cases in particular, is a substance so rich in carbon that it is very near to coal, and so indestructible and impermeable to water, that it contributes more largely than anything else to the mineral. Sir Charles Lyell remarks—"To prevent ourselves, therefore, from hazarding false generalisations, we must ever bear in mind the extreme scantiness of our present information respecting the flora of that peculiar class of stations to which, in the Palæozoic era, the coal-measures probably belonged. I have stated elsewhere my conviction that the plants which produced coal were not drifted from a distance, but nearly all of them grew on the spot where they became fossil. They constituted the vegetation of low regions, chiefly the deltas of large rivers, slightly elevated above the level of the sea, and liable to be submerged beneath the waters of an estuary or sea by the subsidence of the ground to the amount of a few feet. That the areas where the carboniferous[68] deposits accumulated were low, is proved not only by the occasional association of marine remains, but by the enormous thickness of strata of shale and sandstone to which the seams of coal are subordinate. The coal-measures are often thousands of feet, and sometimes two or three miles, in vertical thickness, and they imply that for an indefinite number of ages a great body of water flowed continuously in one direction, carrying down towards a given area the detritus of a large hydrographical basin, draining some large islands or continents, on the margins of which the forests of the coal period grew. If this view be correct, we can know little or nothing of the upland flora of the same era, still less of the contemporaneous plants of the mountainous or alpine regions. If so, this fact may go far to account for the apparent monotony of the vegetation, although its uniform character may doubtless be in part owing to a greater uniformity of climate then prevailing throughout the globe. Mr. Bunbury has successfully pointed out that the peculiarity of the carboniferous climate consisted more in the humidity of the atmosphere and the absence of cold, or rather the equable temperature preserved in the different seasons of the year, than in its tropical heat; but we must still presume that colder climates existed at higher elevations above the sea."
The plants of the coal-measures are evidently terrestrial plants. Brongniart agrees with Lyell in thinking that the layers of coal have in general accumulated in the situation where the plants forming them grew. The remains of these plants covered the soil in the same way as layers of peat, or the vegetable mould of great forests. In a few instances, however, the plants may have been transported from a distance, and drifted into basins. Phillips is disposed to think that this was the general mode of formation of coal-basins. He is led to this conclusion by observing the fragmentary state of the stems and branches, the general absence of roots,[69] and the scattered condition of all the separable organs. Those who support the drift theory, look on the coal plants as having been swept from the land on which they grew by watery currents at different times, and deposited in basins and large sea-estuaries, and sometimes in lakes. The snags in the Mississippi, the St. Lawrence, and other large rivers, are given as instances of a similar drifting process.
The vegetation of the coal epoch seems to resemble most that of islands in the midst of vast oceans, and the prevalence of ferns indicates a climate similar to that of New Zealand in the present day. In speaking of the island vegetation of the coal epoch, Professor Ansted remarks (Ancient World, p. 88)—"The whole of the interior of the islands may have been clothed with thick forests, the dark verdure of which would only be interrupted by the bright green of the swamps in the hollows, or the brown tint of the ferns covering some districts near the coasts. The forests may have been formed by a mixture of several different trees. We would see then, for instance, the lofty and widely-spreading Lepidodendron, its delicate feathery fronds clothing, in rich luxuriance, branches and stems, which are built up, like the trunk of the tree-fern, by successive leafstalks that have one after another dropped away, giving by their decay additional height to the stem, which might at length be mistaken for that of a gigantic pine. There also should we find the Sigillaria, its tapering and elegant form sustained on a large and firm basis—enormous matted roots, almost as large as the trunk itself, being given off in every direction, and shooting out their fibres far into the sand and clay in search of moisture. The stem of this tree would appear like a fluted column, rising simply and gracefully without branches to a great height, and then spreading out a magnificent head of leaves like a noble palm-tree. Other trees, more or less resembling palms, and others like existing firs, also abounded, giving a richness and[70] variety to the scene; while one gigantic species, strikingly resembling the Norfolk Island pine, might be seen towering a hundred feet or more above the rest of the forest, and exhibiting tier after tier of branches richly clothed with its peculiar pointed spear-like leaves, the branches gradually diminishing in size as they approach the apex of a lofty pyramid of vegetation. Tree-ferns also in abundance might there be recognised, occupying a prominent place in the physiognomy of vegetation, and dotted at intervals over the distant plains and valleys, the intermediate spaces being clothed with low vegetation of more humble plants of the same kind. These we may imagine exhibiting their rich crests of numerous fronds, each many feet in length, and produced in such quantity as to rival even the palm-trees in beauty. Besides all these, other lofty trees of that day, whose stems and branches are now called Calamites, existed chiefly in the midst of swamps, and bore their singular branches and leaves aloft with strange and monotonous uniformity. All these trees, and many others that might be associated with them, were, perhaps, girt round with innumerable creepers and parasitic plants, climbing to the topmost branches of the most lofty amongst them, and relieving, in some measure, the dark and gloomy character of the great masses of vegetation."
Hugh Miller remarks—"The sculpturesque character of the nobly-fluted Sigillarias was shared by not a few of its contemporaries. Ulodendrons, with their rectilinear rows of circular scars, and their stems covered with leaf-like carvings, rivalled in effect the ornately relieved torus of a Corinthian column. Favularia, Halonia, many of the Calamites, and all the Lepidodendrons, exhibited the most delicate sculpturing. In walking among the ruins of this ancient flora, the palæontologist almost feels as if he had got among the broken fragments of Italian palaces erected long years ago, when the architecture of Rome was most ornate, and every moulding[71] was roughened with ornament; and in attempting to call up in fancy the old Carboniferous forests, he has to dwell on this peculiar feature as one of the most prominent; and to see in the multitude of trunks darkened above by clouds of foliage that rise upon him in the prospect, the slender columns of an older Alhambra, roughened with arabesque tracery and exquisite filigree work."
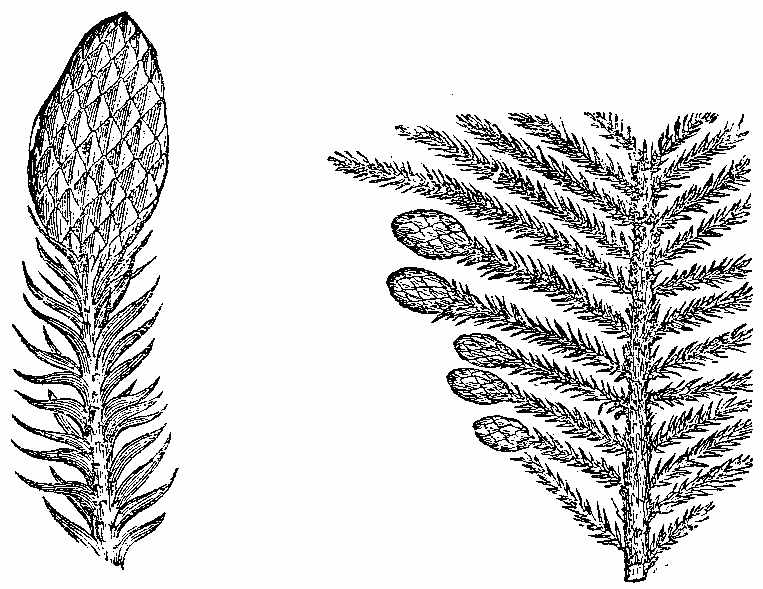
The nature of the vegetation during the Permian period, which is associated with the Carboniferous, under the reign of Acrogens, has been extensively illustrated by Goeppert. Brongniart has enumerated the fossils in three different localities, which he refers doubtfully to this period. 1. The flora of the bituminous slates of Thuringia, composed of Algæ, Ferns, and Coniferæ. 2. Flora of the Permian sandstones of Russia, comprehending Ferns, Equisetaceæ, Lycopodiaceæ, and Noeggerathiæ. 3. Flora of the slaty schists of Lodève, composed of Ferns, Asterophyllites, and Coniferæ. The genera of Ferns here met with are those found in the Carboniferous epoch; the Gymnosperms are chiefly species of Walchia and Noeggerathia (the latter is supposed by Schimper to be a Cycad); Lepidodendron elongatum, Calamites gigas, and Annularia floribunda, are also species of this period. Walchia is a conifer characteristic of the Permian epoch, of which there are eight species described (Figs. 55 and 56). It has a single seed to each scale of the cone, and two kinds of leaves, the one short and imbricated, the other long and spreading. Among the plants of the Permian formation Goeppert enumerates the following:[17]—Equisetites contractus, Calamites Suckowi, C. leioderma, Asterophyllites equisetiformis, A. elatior, Huttonia truncata, H. equisetiformis, many species of Psaronius, one of the filicoid plants, Hymenophyllites complanatus, Sphenopteris crassinervia, Sagenopteris tæniæfolia, Neuropteris imbricata, and many other species of these[72] genera; several species of Odontopteris, Callipteris, Cyclopteris, Dioonopteris, Cyatheites, Alethopteris, Noeggerathia, Cordaites, Anthodiopsis, Dictyothalamus, Calamodendron, Arthropitys; besides species of Sigillaria, Stigmaria, and Lepidodendron. Various fruits are also mentioned, under the names of Rhabdocarpum, Cardiocarpum, Acanthocarpum, Trigonocarpum, and Lepidostrobus.

The Gymnospermous plants of the present day are included in two natural orders, Coniferæ and Cycadaceæ. Under Coniferæ are enumerated the various species of Pine (Fig. 57), Spruce (Fig. 58), Larch (Fig. 59), Cedar (Fig. 60), Eutassa,[73] Araucaria (Fig. 61), Sequoia, Cryptomeria, Taxodium, Cypress, Juniper (Fig. 70), Salisburia, Dacrydium, Yew (Fig. 71), etc.
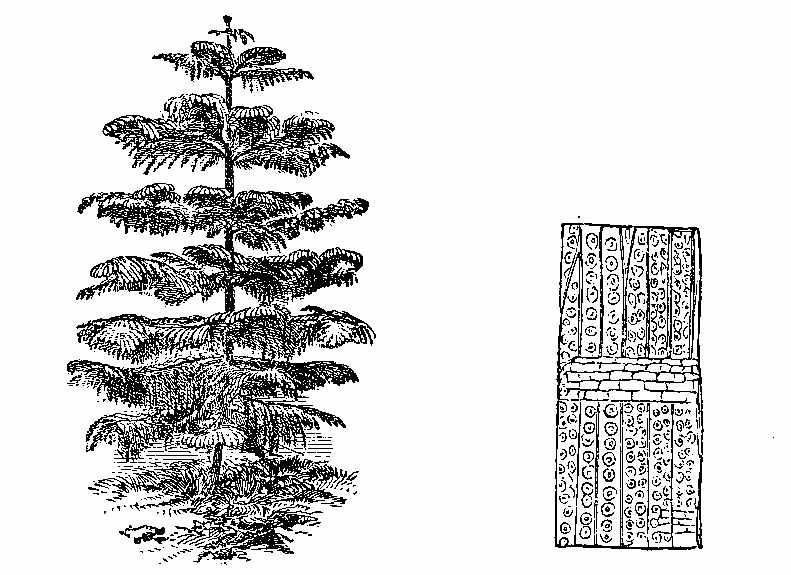
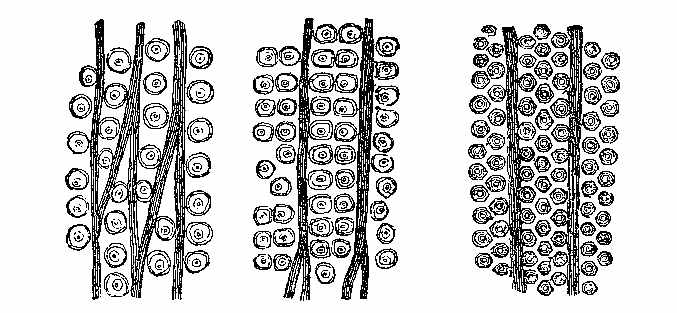
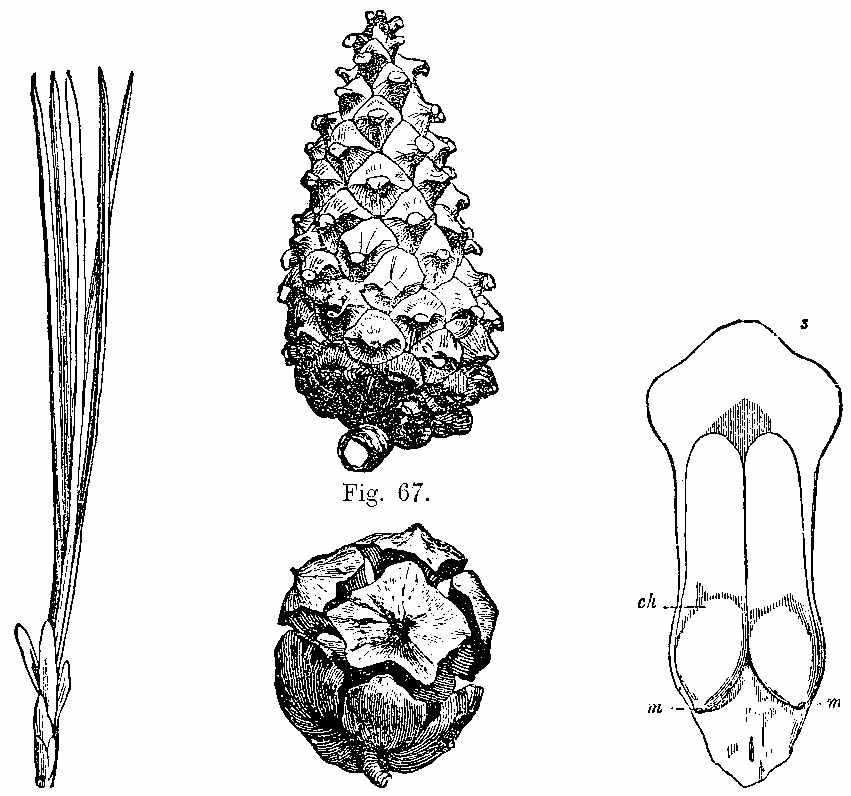
The Coniferæ of the present day are distinguished as resinous trees or shrubs with punctated woody tissue (Figs. 62, 63, 64, 65), linear acerose or lanceolate parallel-veined leaves, sometimes clustered, and having a membranous sheath[75] at the base (Fig. 66). Male flowers in deciduous catkins; female flowers in cones (Figs. 67, 68). The seeds are considered by most botanists as being naked, i.e. not contained in a true pistil (Fig. 69). Some of the conifers have a succulent cone, as the juniper (Fig. 70), and the yew (Figs. 71-73) has a succulent mass covering a single naked seed (Fig. 73). The yew also has its pleurenchyma marked both with punctations and spiral fibres. The arrangement of the punctations in the Coniferæ gives characters which enable us to classify the woods into groups that have some relation to the[76] genera established from the reproductive organs (see Figs. 62-65).

The natural order Cycadaceæ is not so largely represented at the present day as it was during the Mesozoic epoch. Among the genera of the present day are Cycas (Fig. 74), Zamia, Macrozamia, Encephalartos (Fig. 75), Dion, Stangeria, etc. They are small palm-like trees or shrubs, with unbranched stems, occasionally dichotomous, marked with leaf-scars, and having large medullary rays along with pitted woody tissue. The leaves are pinnate, except in Bowenia, which has a bipinnate leaf. Males in cones. Females consisting of naked ovules on the edges of altered leaves, or on the inferior surface of the peltate apex of scales.[18]

In this reign the Acrogenous species are less numerous; the Gymnosperms almost equal them in number, and ordinarily surpass them in frequency. There are two periods in this reign, one in which Coniferæ predominate, while Cycadaceæ scarcely appear; and another in which the latter family preponderates as regards the number of species, and the frequency and variety of generic forms. Cycadaceæ occupied a more important place in the ancient than in the present vegetable world. They extend more or less from the Trias formation up to the Tertiary. They are rare in the Grès bigarré or lower strata of the Triassic system. They attain their maximum in the Lias and Oolite, in each of which upwards of 40 species have been enumerated, and they disappear in the Tertiary formations. Schimper describes 13 genera of fossil Zamiæ, and about 20 Cycadeæ. He thinks that Trigonocarpum (15 species), Rhabdocarpum (24 species),[78] Cardiocarpum (21 species), and Carpolithes (9 species), are all fruits of Cycadeæ. Many supposed fossil Cycads are looked upon by Carruthers as Coniferæ. Zamia macrocephala, or Zamites macrocephalus, or Zamiostrobus macrocephalus, is called by him Pinites macrocephalus; Zamia ovata, or Zamites ovatus, or Zamiostrobus ovatus, is Pinites ovatus; Zamia Sussexiensis is Pinites Sussexiensis. Among other species of Pinites noticed by Carruthers are Pinites oblongus, P. Benstedi, P. Dunkeri, P. Mantellii, P. patens, P. Fittoni, P. elongatus. It is important to notice that in an existing Cycad called Stangeria paradoxa the veins of the pinnæ rise from a true midrib and fork, characters which render untenable the distinction usually relied upon between the foliage of Ferns and Cycads.
In Brongniart's Vosgesian period, the Grès bigarré, or the Red Sandstones and Conglomerates of the Triassic system, there is a change in the flora. Sigillarias and Lepidodendrons disappear, and in their place we meet with Gymnosperms, belonging to the genera Voltzia, Haidingera, Zamites, Ctenis,[79] Æthophyllum, and Schizoneura (Fig. 76). The genus Voltzia is confined to the Trias, and though a true conifer, it is not easy to correlate it with any living form. It is apparently Abietineous, having two seeds to each scale, but they are placed on the dilated upper portion of the scale. The leaves are of two kinds, the one broad and short, and the other at the tops of the branches long and linear. Species of Neuropteris, Pecopteris, and other acrogenous coal genera are still found, along with species of Anomopteris and Crematopteris—peculiar Fern-forms, which are not found in later formations. Stems of arborescent Ferns are more frequent than in the next period.
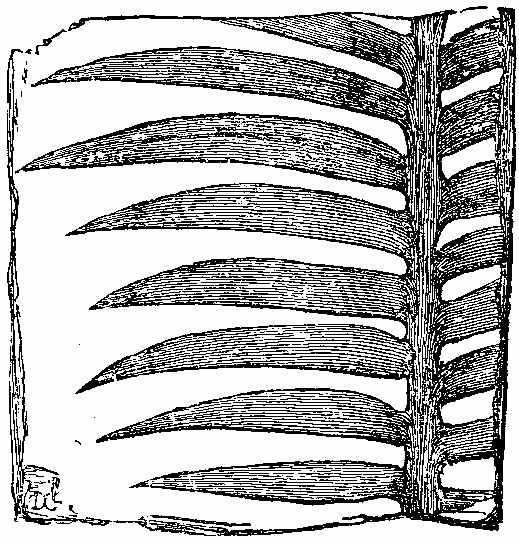
The Jurassic period of Brongniart embraces the Keupric period or variegated marls of the Triassic system, the Liassic epoch, the Oolitic and the Wealden. The flora of the Keupric epoch differs from that of the Grès bigarré of the Vosges. The Acrogens are changed as regards species, and frequently in their genera. Thus we have the genera Camptopteris, Sagenopteris, and Equisetum. Among Gymnosperms, the genera Pterophyllum and Taxodites occur.
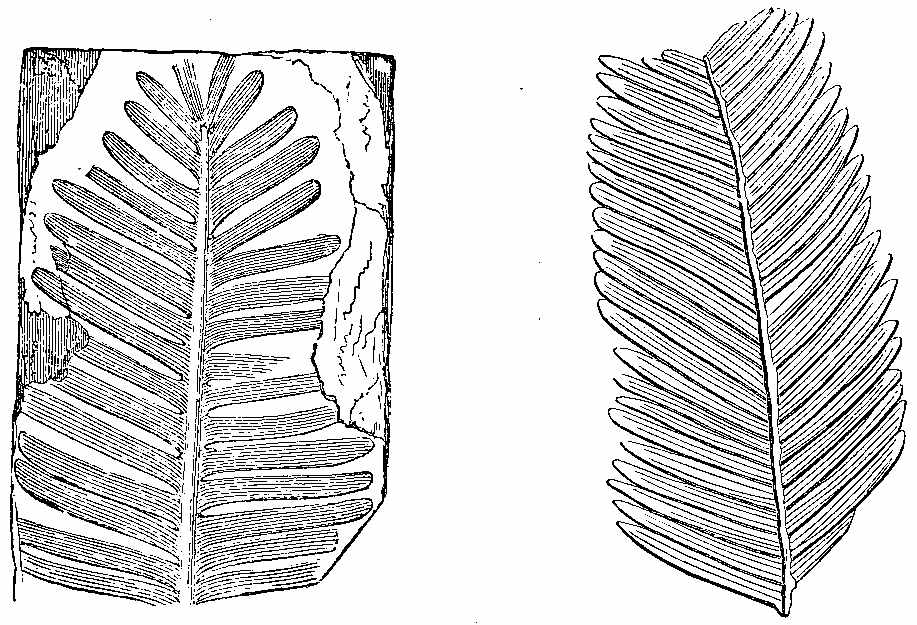
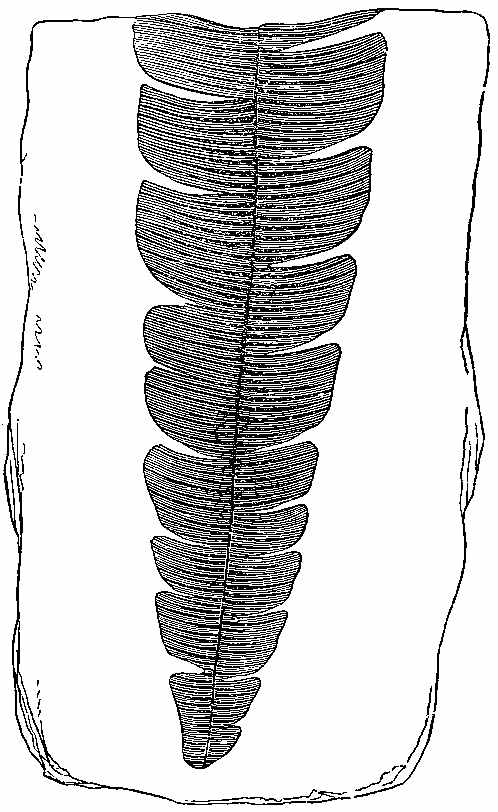
In the Lias the essential characters of the flora are the predominance of Cycadaceæ, in the form of species of Cycadites, Otozamites, Zamites (Fig. 77), Ctenis, Pterophyllum (Fig. 78), and Nilssonia (Fig. 79), Palæozamia (Fig. 80), and the existence among the Ferns of[80] many genera with reticulated venation, such as Camptopteris and Thaumatopteris, some of which began to appear at the Keupric epoch. Coniferous genera, as Brachyphyllum (Fig. 81), Taxodites, Palissya, and Peuce, are found. In the Lias near Cromarty, Miller states that he found a cone with long bracts like those of Pinus bracteata.
In the Oolitic epoch the flora consists of numerous Cycadaceæ and Coniferæ, some of them having pecu[81]liar forms. Its distinctive characters are, the rarity of Ferns with reticulated venation, which are so numerous in the Lias, the frequency of the Cycadaceous genera Otozamites and Zamites, which are most analogous to those now existing; the occurrence of a remarkable group presenting very anomalous structure in their organs of reproduction, to which Carruthers has given the name of Williamsonia; and the diminution of Ctenis, Pterophyllum, Palæozamia, and Nilssonia, genera far removed from the living kinds; and lastly, the greater frequency of the coniferous genera, Brachyphyllum and Thuites, which are much more rare in the Lias. In the Scottish Oolite at[82] Helmsdale, Miller detected about 60 species of plants, including Cycadaceæ and Coniferæ, with detached cones, and Fern-forms resembling Scolopendrium. He also discovered a species of Equisetum, and what he supposed to be a Calamite.
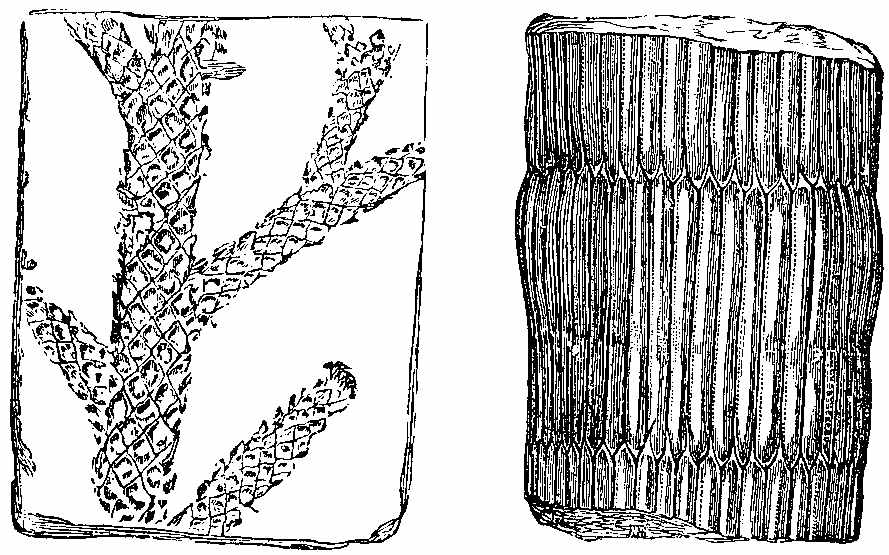
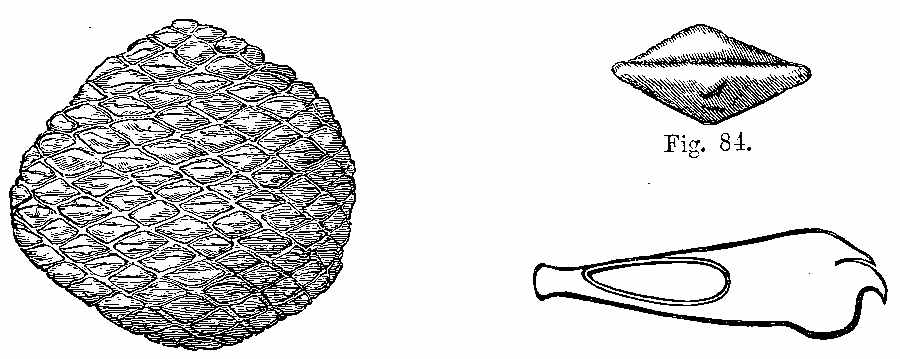

There is an absence of true coal-fields in the secondary formations generally; but in some of the Oolitic series, as in the lower Oolite at Brora, in Sutherlandshire, and in the north-east of Yorkshire, and the Kimmeridge clay of the upper Oolite, near Weymouth, there are considerable deposits of carbonaceous matter, sometimes forming seams of coal which have been worked for economic purposes.[19] Some suppose that the Brora coal was formed chiefly by Equisetum columnare (Fig. 82). In the sandstones and shales of the Oolitic series, especially in the lower Oolite of the north of England, as at Whitby and Scarborough, as well as in Stonesfield slate, the Portland Crag of the middle, and the Portland beds of the upper Oolite, numerous fossil plants are found. Peuce Lindleyana is one of the Coniferæ of the lower Oolite. Beania (Plate II. Fig. 2) is a Cycadaceous[83] fossil from the Oolite of Yorkshire (Carruthers, Geol. Mag. vi. 91). Araucarites sphærocarpus (Figs. 83, 84, 85) is found in the inferior Oolite, and separate scales of Araucarian fruits occur in the Oolitic shales of Yorkshire (Araucarites Phillipsii, Plate II. Fig. 11), and in the "slate" at Stonesfield (A. Brodiei, Plate II. Fig. 10). The upper Oolite at Portland contains an interesting bed, about a foot in thickness, of a dark brown substance. This is the Dirt-bed (Fig. 86) made up of black loam, which, during the Purbeck period, formed a surface soil which was penetrated by the roots of trees, fragments of whose stems are now found in it fossilised. These consist of an assemblage of silicified stumps or stools of large trees, from 1-3 feet high, standing in their original position, with the roots remaining attached to them, and still penetrating the earth in which they grew. Besides the erect trunks many stems have been broken and thrown down, and are buried in a horizontal position in the bed. They belong to Coniferæ and Cycadaceæ. One of these is Mantellia nidiformis, shown in Fig. 87. Carpolithes conicus and C. Bucklandi are fruits found in the Oolite. Some look upon them as fruits of palms.
Several species of Pandanaceous fruits have been found in Oolitic strata. Buckland described one of them as Podocarya, which is remarkable, as it consists of a single but many-seeded drupe. To another form, more nearly allied to the existing plants, Carruthers has given the name Kaidacarpum, and has described three species. These fruits are made up of a large number of single-seeded drupes. The species figured (Fig. 88) is from the great Oolite, near Northampton. In Fig. 89 a representation is given of one of the Pandanaceæ, the screw-pines of the present day.
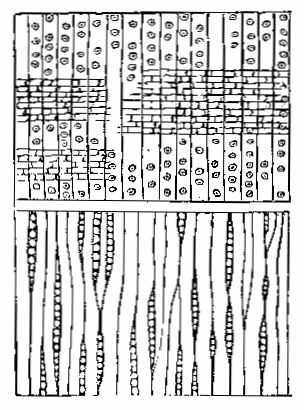
The flora of the Wealden epoch is characterised in the south of England by the abundance of the fern called Lonchopteris Mantellii, and in Germany by the predominance of the Conifer denominated Abietites Linkii (Fig. 90), and the presence of Araucarites Pippingfordensis, as well as by numerous Cycadaceæ, such as species of Cycadites, Zamites, Pterophyllum, Mantellia, Bucklandia, and a remarkable genus having a fleshy fruit, and related to the ordinary Cycadaceæ as Taxus is to the other Coniferæ, which has been fully described in[85] the Linn. Trans., under the name of Bennettites (Plate II. Fig. 3). In the Wealden at Brook Point, Isle of Wight, Cycads have been detected allied to Encephalartos. The fruits of them are described by Carruthers as Cycadeostrobus. He describes the following species:—Cycadeostrobus ovatus (Plate II. Fig. 1), C. truncatus, C. tumidus, C. elegans, C. Walkeri, C. sphæricus, in the Oxford clay of Wiltshire; C. primævus in the inferior Oolite at Burcott Wood and Livingston, and C. Brunonis. Mantell states that he has found 40 or 50 fossil cones in the Wealden of England; they belong either to the genus Cycadeostrobus or to the pines mentioned below as occurring in the Wealden. The Wealden fresh-water formation terminates the reign of Gymnosperms. Carruthers gives the following list of the remains of Coniferæ which have been found in the secondary strata of Britain, excluding the Trias:—
Upper Chalk.—Wood in flint nodules.
Upper Greensand.—Foliage and cone of Sequoiites Woodwardii; cone of Pinites oblongus.
Gault.—Cones of Pinites gracilis and P. hexagonus, Sequoiites Gardneri and S. ovalis.
Lower Greensand.—Water-worn and bored pieces of wood; cones of Pinites Benstedi, P. Sussexiensis, and P. Leckenbyi.
Wealden.—Driftwood, foliage of Abietites Linkii; cones of Pinites Dunkeri, P. Mantellii, P. patens, and P. Fittoni, and of Araucaria Pippingfordensis; foliage (and drupes?) of Thuites Kurrianus.
Purbeck.—Fossil forest in situ at Isle of Portland; cone "nearly related to Araucaria excelsa" in the Dirt-bed.
Portland Stone.—Driftwood Araucarites.
Kimmeridge Clay.—Cone of Pinites depressus.
Oxford Clay.—Driftwood and foliage of Araucarites.
Great Oolite.—Driftwood of Araucarites; foliage of Thuites acutifolius, T. articulatus, T. cupressiformis, T. divaricatus, and T. expansus, and of Taxites podocarpoides; detached cones at Helmsdale, Sutherland.
Inferior Oolite.—Wood of Peuce Eggensis (Tertiary according to Geikie); foliage of Brachyphyllum mammillare, Cryptomerites? divaricatus, and Palissya? Williamsonis; cones of Araucarites sphærocarpus, A. Brodiei, and A. Phillipsii. Pinites primæva (Lindl. and Hutt.) is a Cycadean fruit.
Lias.—Wood of Pinites Huttonianus and P. Lindleyanus; foliage of Araucaria peregrina and Cupressus latifolia; cone of Pinites elongatus, and "cone with long bracts like those of Pinus bracteata," from Cromarty.
Carruthers gives the following arrangement of fossil Cycadaceæ in the Transactions of the Linn. Soc. vol. xxvi.—Firstly, the Cycadeæ: including the genus Bucklandia, Presl; and species B. anomala, B. Mantellii, B. squamosa, B. Milleriana—the two first-named species being from the Wealden, and the two last-named from the Oolite. Secondly, the Zamieæ: including the genus Yatesia, Carr.; and species Y. Morrisi, Lower Cretaceous; Y. gracilis, Lias; Y. crassa, M. Oolite; Y. Joassiana, M. Oolite; the genus Fittonia, Carr., and species F. squamata, U. Cretaceous; the genus Crossozamia, Pomel, and species C. Moreaui, Pomel, Jurassic, and C. Buvignieri, Pomel, Jurassic—both from St. Michel, France. Thirdly, the Williamsonieæ: including the genus Williamsonia, Carr.; and species W. gigas, W. pecten, W. hastula, all from the inferior Oolite. Fourthly, the Bennettiteæ: including the genus Bennettites, Carr., and species B. Saxbyanus, Wealden; B. Gibsonianus, Lr. Greensand; B. maximus, Wealden; B. Portlandicus, Lr. Purbeck; and B. Peachianus, M. Oolite; the genus Mantellia, Brong., and species M. nidiformis, M. intermedia, M. microphylla, from the Lr. Purbeck; and M. inclusa, from the Lr. Cretaceous; the genus Raumeria, Goeppert, and species R. Reichenbachiana, from Galicia, and R. Schulziana from Silesia.
(INCLUDING THE CRETACEOUS EPOCH).
This reign is characterised by the appearance of Angiospermous Dicotyledons, plants which constitute more than three-fourths of the species of the existing flowering plants of the globe, and which appear to have acquired the predominance from the commencement of the Tertiary epoch. They are plants with seeds contained in seed-vessels, and each seed with two cotyledons. These plants, however, appear even at the beginning of the Cretaceous period. In this reign, therefore, Brongniart includes the upper Secondary period, or the Cretaceous system, and all the Tertiary period. The Cretaceous may be considered as a sort of transition period between the reign of Gymnosperms and Angiosperms.
The Chalk flora is characterised by the Gymnospermous almost equalling the Angiospermous Dicotyledons, and by the existence of a considerable number of Cycadaceæ, which do not appear in the Tertiary period. The genus Credneria is one of the characteristic forms. In this period we find Algæ represented by Cystoseirites, Confervites, Sargassites, and Chondrites; Ferns by peculiar species of Pecopteris and Protopteris; Naiadaceæ by Zosterites; Palms, by Flabellaria and Palmacites; Cycadaceæ by Cycadites, Zamites, Microzamia, Fittonia, and Bennettites; Coniferæ, by Brachyphyllum, Widdringtonites, Cryptomeria, Abietites, Pinites, Cunninghamites, Dammarites, Araucarites; and Angiospermous Dicotyledons, by Comptonites, Alnites, Carpinites, Salicites, Acerites, Juglandites, and Credneria. At the base of the Tertiary period there are deposits of Algæ of a very peculiar form, belonging to the genera Chondrites and Munsteria. No land plants have been found mingled with these marine species.
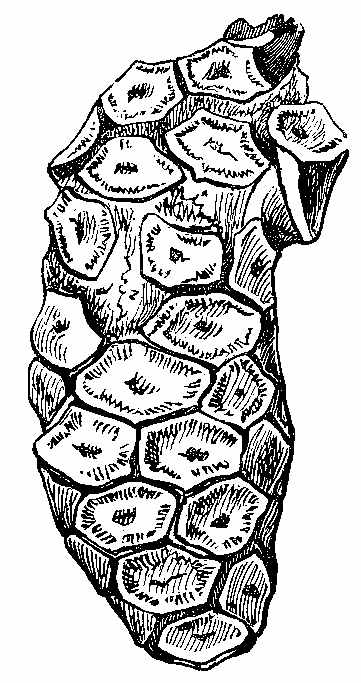
In the Gault, near Folkestone, an interesting association of coniferous fruits has been found, consisting of two species of Sequoia, along with two of Pinus. The pines belong to the same group as those which now grow with the Wellingtonias in California, showing the remarkable fact that the coniferous vegetation of the high lands of the Upper Cretaceous period had a facies similar to that now existing in the mountains on the west of North America. We figure both the species of Sequoiites—viz. S. ovalis (Fig. 91), a large cone, and S. Gardneri (Plate II. Fig. 7). In the present day there are two species of the genus Sequoia—viz. S. gigantea (Wellingtonia gigantea) and S. sempervirens.[20] In the Lower Greensand a remarkably fine cone belonging to the same group as the Cedar has been found. This is the Pinites Leckenbyi (Plate II. Fig. 4). A section exhibits the seeds in their true position, some of which are preserved so as to exhibit the form and position of the embryo.
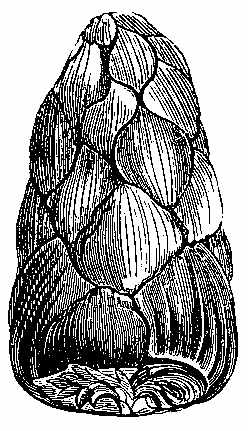
The Tertiary period is characterised by the abundance of Angiospermous Dicotyledons and of Monocotyledons, more especially of Palms. By this it is distinguished from the more ancient periods. Angiosperms at this period greatly exceed Gymnosperms. Cycadaceæ are very rare, if not completely wanting, in the European Tertiary strata, and the Coniferæ belong to genera of the temperate regions. In the lower Tertiaries Carruthers has found a fossil Osmunda, and the existence of a group of Pines having cones with a very thick apophysis. From their remarkable external aspect,[89] these cones had been considered to be Cycadean, but their internal structure indicates that they are coniferous. Pinites ovatus is one of these cones (Fig. 92). The Cupressineæ are found in the Tertiary beds only. Taxodieæ are represented by Sequoiites (Plate II. Fig. 7) in the Cretaceous and Eocene strata. Peuce australis of Van Diemen's Land and P. Pritchardi of Ireland are Tertiary plants. The Peuce of Eigg (P. Eggensis), according to Geikie, is also Tertiary, and not Oolitic. Isoetes is mentioned by Schimper as a Tertiary genus. Although the vegetation throughout the whole of the Tertiary period presents pretty uniform characters, still there are notable differences in the generic and specific forms, and in the predominance of certain orders at different epochs. Brongniart does not entirely agree with Unger as to these epochs. Many of the formations classified by Unger in the Miocene division he refers with Raulin to the Pliocene. He divides the Tertiary period, as regards plants, into the Eocene, Miocene, and Pliocene epochs, and gives the following comparative results from an examination of their floras:—
| Classes and Sub-Classes. | Eocene Epoch. | Miocene Epoch. | Pliocene Epoch. |
| Thallogenæ | 16 | 6 | 6 |
| Acrogenæ | 17 | 4 | 7 |
| Monocotyledones | 33 | 26 | 4 |
| Dicotyledones— | |||
| Gymnospermæ | 40 | 19 | 31 |
| Angiospermæ | 103 | 78 | 164 |
| 209 | 133 | 212 | |
In the Eocene formation the fossil fruits of the Isle of[90] Sheppey increase the number of Phanerogamous plants, only a small proportion of which have as yet been described. This is an exceptional locality, and the deposit in which the fruits occur is probably the silt found at the mouth of a large river which flowed, like the Nile, from tropical regions towards the north. The number of plants as given by Brongniart is much smaller than that mentioned by Unger (p. 23). The latter includes in his enumeration a considerable amount of uncertain species.
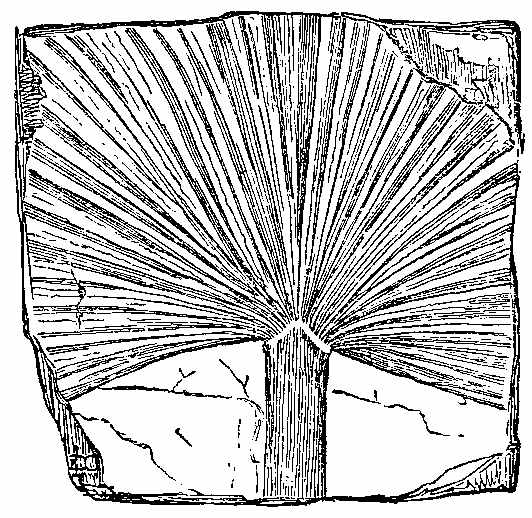
The Eocene epoch in general is characterised by the predominance of Algæ and marine Naiadaceæ, such as Caulinites and Zosterites; by numerous Coniferæ, the greater part resembling existing genera among the Cupressineæ, and appearing in the form of Juniperites, Thuites, Cupressinites (Plate II. Figs. 8, 9), Callitrites, Frenelites, and Solenostrobus; by the existence of a number of extra-European forms, especially of fruits, such as Nipadites, Leguminosites, Cucumites, and Hightea; and by the presence of some large species of Palm belonging to the genera Flabellaria and Palmacites (Fig. 93).
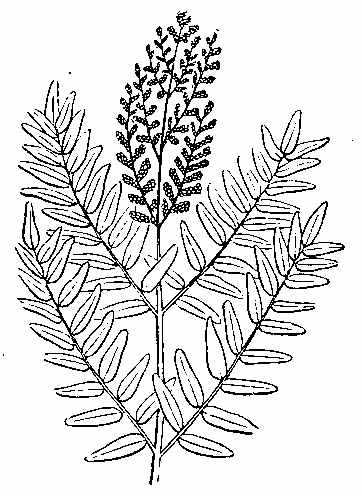
Unger says that the Eocene flora has resembled in many respects that of the present Australian vegetation. He gives the following genera as occurring at the Eocene epoch:—Araucaria, Podocarpus, Libocedrus, Callitris, Casuarina, Pterocarpus, Drepanocarpus, Centrolobium, Dalbergia, Cassia, Cæsalpinia, Bauhinia, Copaifera, Entada, Acacia, Mimosa, Inga. (Seemann's Journal of Bot. vol. iii. p. 43.) Amber is considered[91] to be the produce of many Coniferæ of this epoch, such as Peuce succinifera or Pinites succinifera, and Pinus Rinkianus. It occurs in East Prussia in great quantity, and it is said that many pieces of fossil wood occur there, which, when moderately heated, give out a decided smell of amber. Connected with these beds are found cones belonging to Pinites sylvestrina and P. Pumilio-miocena, species nearly allied to the living species; others to Pinites Thomasianus and P. brachylepis. Goeppert contrasts the present flora of Germany and that of the Amber epoch as follows:—
| German Flora. | Amber Flora. | |
| Cryptogameæ | 6800 | 60 |
| Phanerogameæ | 3454 | 102 |
| and gives the following specimens of two of the orders:— | ||
| Cupuliferæ | 12 | 10 |
| Ericaceæ | 23 | 24 |
(See remarks by Goeppert on the Amber Flora, etc., Edin. N. Phil. Journ. lvi. 368; and Quart. Journ. Geol. Soc. x. 37.) In the lower Eocene of Herne Bay, Carruthers noticed a fern like Osmunda (Fig. 94), which he calls Osmundites Dowkeri (Plate I. Figs. 8, 9). This specimen was silicified; starch grains contained in its cells, and the mycelium of a parasitic fungus traversing some of them, were perfectly preserved. Berkeley has detected in amber fossil fungi, which he has named Penicillium curtipes, Brachycladium Thomasinum, and Streptothrix spiralis.[21] Some Characeæ are also met with, as[92] Chara medicaginula and C. prisca, with a fossil called Gyrogonites, the nucule or the fructification of these plants. Carpolithes ovatus, a minute seed-vessel, occurs in the Eocene beds of Lewisham. Another small fruit, of a similar nature, called Folliculites minutulus, occurs in the Bovey Tracey coal, which belongs to the Tertiary beds.
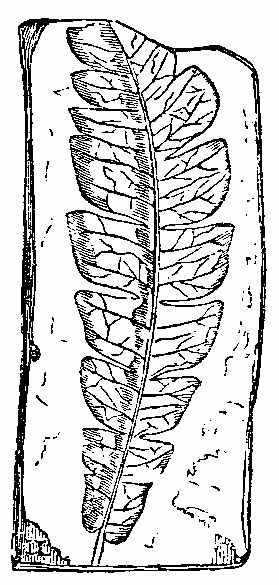
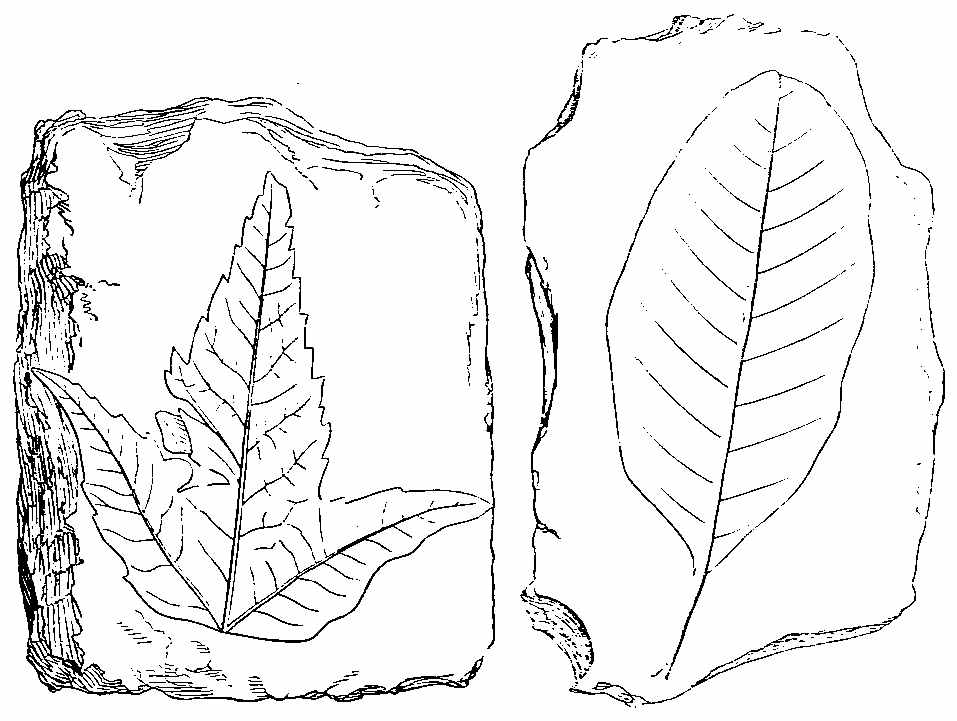

The most striking characters of the Miocene epoch consist in the mixture of exotic forms of warm regions with those of temperate climates. Unger says that it resembles that of the southern part of North America. Thus we meet with Palms, such as species of Flabellaria and Phœnicites, a kind of Bamboo called Bambusium sepultum; Lauraceæ, as Daphnogene and Laurus; Combretaceæ, as Getonia and Terminalia; Leguminosæ, as Phaseolites, Desmodophyllum, Dolichites, Erythrina, Bauhinia, Mimosites, and Acacia—all plants having their living representatives in warm climates; Echitonium, Plumiera, and other Apocynaceæ of equatorial regions; Comptonia (Fig. 95), a Proteaceous genus, and Steinhauera, a Cinchonaceous genus; mingled with species of Acer (Fig. 96), Ulmus (Fig. 97), Rhamnus (Fig. 98); and Amentiferous forms, such as Myrica, Betula, Alnus (Fig. 99), Quercus, Fagus, Carpinus, all belonging to temperate and cold climates. The statements as to the occurrence of Pinus sylvestris and Betula alba among the Miocene fossils have not been founded on complete data. It is by no means easy, even in the present day, to distinguish fragments of dried specimens of Pinus Pumilio from those of P. sylvestris, and from a great many[93] other Pines. The difficulty is still greater in fossils (Hook. Kew Journ. v. 413). There are a very small number of plants belonging to orders with gamopetalous corollas. In the Miocene formation of Lough Neagh in Ireland, and of Mull in Scotland, silicified trunks of considerable size have been found. The Irish silicified wood has been denominated Cupressoxylon Pritchardi from its apparent resemblance to the Cypress. As connected with the Miocene epoch, we may notice the leaf-beds found at Ardtun, in the island of Mull,[94] by the Duke of Argyll.[22] Above and below these beds basalt occurs, and there are peculiar tuff-beds alternating with the leafy deposits. These tuff-beds were formed by the deposit of volcanic dust in pools probably of fresh water. They contain fragments of chalk and flint. The leaves are those of plants allied to the Yew, Rhamnus, Plane, and Alder, along with the fronds of a peculiar Fern, and the stems of an Equisetum. The genera are Taxites or Taxodites (Fig. 100), Rhamnites (Fig. 101), Platanites, Alnites,[95] Filicites, and Equisetum (Fig. 102). In the leaf-beds at Bournemouth Mr. Wanklyn detected several ferns. One is a species of Didymosorus, and shows distinct venation and fructification. Fossilised wood was found in the Arctic Regions by Captain M'Clure. At the N.W. of Banks Land he found trees with trunks 1 foot 7 inches in diameter.


[96] Dr. Oswald Heer[23] has examined the plants preserved in the lignite beds of Bovey Tracey, in Devonshire, and he finds that they belong to the Miocene formation. There is a remarkable coincidence between this and several of the continental fossil floras, such as those of Salzhauser in the Wetterau, Manosque in Provence, and of some parts of Switzerland. Bovey Tracey has no species in common with Iceland, although the Tertiary flora of Iceland belongs to the same period. Two of its species (Corylus MacQuarrii and Platanus aceroides) have been found in the Miocene of Ardtun Head. Even the genera are distinct, with the exception of Sequoia and Quercus. The Bovey Tracey flora has a much more southern character, corresponding entirely with that of the Lower Miocene of Switzerland. It contains three species of Cinnamon, one Laurel, evergreen Figs, one Palm, and large Ferns, thus manifesting a subtropical climate. One of the most important plants is Sequoia Couttsiæ, a Conifer which supplies a link between S. Langsdorfii and S. Sternbergi, the widely-distributed representatives of S. sempervirens and S. gigantea (Wellingtonia), which are Californian trees. Among other characteristic plants may be mentioned Cinnamomum lanceolatum and C. Scheuchzeri; Quercus Lyellii, an evergreen oak; species of evergreen fig (Ficus Falconeri and F. Pengellii), Palmacites Dæmonorops, a prickly twining Rotang-palm; species of Vine (Vitis Hookeri and V. Britannica); Pecopteris lignitum, a large tree-fern; species of Nyssa, at present confined to North America. Among other plants recorded by Heer in his paper are the following:—Laurus primigenia, Daphnogene Ungeri, species of Dryandroides, Andromeda, Vaccinium acheronticum, Echitonium cuspidatum,[97] Gardenia Wetzleri, species of Anona, Nymphæa Doris, Carpolithes Websteri, C. Boveyanus, and other species. In the post-tertiary white clay of Bovey Tracey, Salix cinerea, and a species allied to S. repens, as well as Betula nana, are found.
The Arctic fossil flora (Miocene), according to Heer, amounts to 162 species: Cryptogamia, 18 species, of which 9 are large ferns; Phanerogamia, Coniferæ, 31; Monocotyledons, 14; Dicotyledons, 99. Among the Coniferæ are—Pinus M'Clurii, Sequoia Langsdorffii, Sternbergi, and Couttsiæ, Taxodium dubium, Glyptostrobus europæus, Thujopsis europæa. Among leafy trees are—Fagus Deucalionis, Quercus Olafseni, Platanus aceroides, willows, beeches, Acer, Otopteryx, tulip-tree, walnuts, Magnolia Inglefieldi, Prunus Scottii, Tilia Malmgreni, Corylus M'Quarrii, Alnus Kefersteinii, Daphnogene Kannii, probably one of the Lauraceæ; and among Proteaceæ, MacClintockia? and Hakea. In Greenland are found species of Rhamnus, Paliurus, Cornus, Ilex, Cratægus, Andromeda, Myrica, Ivy, and Vine. From the flora of Spitzbergen, in the Miocene epoch, we may conclude that under 79° N. lat. the mean temperature of the year may have been 41° Fahr., while at the same epoch that of Switzerland was 69°·8 Fahr.; judging from the analogy of floras, it appears that the mean temperature has fallen 6°·9. From this it follows that at Spitzbergen, at 78° N. lat., the mean temperature was perhaps 41°·9 Fahr. In Greenland, at 70°, it would be 49°·1 Fahr., and in Iceland and on the Mackenzie, in lat. 65°, it would be 52°·7 Fahr. At the Miocene epoch the temperature seems to have been much more uniform, the mean heat diminishing much more gradually in proportion as the pole was approached. The isothermal line of 32° Fahr. might have fallen upon the pole, while now it is situated under 58° N. (See Heer's conclusions as to changes of temperature depending on proportion of sea and[98] land, eccentricity of the earth, and the earth moving through warm and cold spaces in the universe—Ann. Nat. Hist. 4th ser. i. 66.)
In speaking of the Polar flora of former epochs, Heer says that every plant executes a slow and continuous migration. These migrations, the starting-point of which is the distant past, are recorded in the rocks; and the interweaving of the carpets of flowers which adorn our present creation retraces them for us in its turn. For the vegetation of the present day is closely connected with that of preceding epochs; and throughout all these vegetable creations reigns one thought, which not only reveals itself around us by thousands upon thousands of images, but strikes us everywhere in the icy regions of the extreme north. Organic nature may become impoverished there, and even disappear when a cold mantle of ice extends over the whole earth; but where the flowers die the rocks speak, and relate the marvels of creation; they tell us that even in the most distant countries, and in the remotest parts, nature was governed by the same laws and the same harmony as immediately around us.[24]
The flora of the Pliocene epoch has a great analogy to that of the temperate regions of Europe, North America, and Japan. We meet with Coniferæ, Amentiferæ, Rosaceæ, Leguminosæ, Rhamnaceæ, Aceraceæ, Aquifoliaceæ, Ericaceæ, and many other orders. There is a small number of Dicotyledons with gamopetalous corollas. The twenty species with such corollas recognised by Brongniart are referred to the Hypogynous Gamopetalous group of Exogens, which in the general organisation of the flowers approach nearest to Dialypetalæ. In this flora there is the predominance of Dicotyledons in number and variety; there are few Monocotyledons.[99] No species appear to be identical, at least with the plants which now grow in Europe. Thus the flora of Europe, even at the most recent geological epoch of the Tertiary period, was very different from the European flora of the present day.
Taking the natural orders which have at least four representatives, Raulin[25] gives the following statement as to the Tertiary flora of central Europe. The Eocene flora of Europe is composed of 128 species, of which 115 belong to Algæ, Characeæ, Pandanaceæ, Palmæ, Naiadaceæ, Malvaceæ, Sapindaceæ, Proteaceæ, Papilionaceæ, and Cupressineæ. The Miocene flora has 112 species, of which 69 belong to Algæ, Palmæ, Naiadaceæ, Apocynaceæ, Aceraceæ, Lauraceæ, Papilionaceæ, Platanaceæ, Quercineæ, Myricaceæ, and Abietineæ. The Pliocene flora has 258 species, of which 226 belong to Algæ, Fungi, Musci, Filices, Palmæ, Ericaceæ, Aquifoliaceæ, Aceraceæ, Ulmaceæ, Rhamnaceæ, Papilionaceæ, Juglandaceæ, Salicaceæ, Quercineæ, Betulaceæ, Taxaceæ, Cupressineæ, and Abietineæ. The Eocene species are included in genera which belong at the present day to inter-tropical regions, comprising in them India and the Asiatic islands of Australia. Some are peculiar to the Mediterranean region. The aquatic plants, which form almost one-third of the flora, belong to genera now peculiar to the temperate regions of Europe and of North America, or occurring everywhere. The Miocene species belong to genera, of which several are found in India, tropical America, and the other inter-tropical regions, but which for the most part inhabit the sub-tropical and temperate regions, including the United States. Some of the genera are peculiar to the temperate regions. The aquatic genera, poor in species, occur everywhere, or else solely in the temperate regions. The Pliocene species belong to genera which almost all inhabit the temperate regions, either of the old[100] continent or of the United States. A few only are of genera existing in India, Japan, and the north of Africa. These various floras, which present successively the character of those of inter-tropical, sub-tropical, and temperate regions, seem to indicate that central Europe has, since the commencement of the Tertiary period, been subjected, during the succession of time, to the influence of these three different temperatures. It would appear, then, Raulin remarks, that the climate of Europe has during the Tertiary period gradually become more temperate.
Brown coal occurs in the upper Tertiary beds, and in it vegetable structure is easily seen under the microscope. Goeppert, on examining the brown coal deposits of northern Germany and the Rhine, finds that Coniferæ predominate in a remarkable degree; among 300 specimens of bituminous wood collected in the Silesian brown coal deposits alone, only a very few other kinds of Exogenous wood occur. This seems remarkable, inasmuch as in the clays of the brown coal formation in many other places leaves of deciduous Dicotyledonous trees have been found; and yet the stems on which we may suppose them to have grown are wanting. The leaves have been floated away from the place where they grew by a current of water which was not powerful enough to transport the stems. The coniferous plants of these brown coal deposits belong to Taxineæ and Cupressineæ chiefly; among the plants are Pinites protolarix and Taxites Ayckii. Many of the Coniferæ exhibit highly compressed, very narrow annual rings, such as occur in Coniferæ of northern latitudes. Goeppert has described a trunk, or rather the lower end of a trunk, of Pinites protolarix, discovered in 1849 in the brown coal of Laasan in Silesia. It was found in a nearly perpendicular position, and measured more than 32 feet in circumference. Sixteen vast roots ran out almost at right angles from the base of the trunk, of which about four feet stood up perfect[101] in form, but stripped of bark. Unfortunately the interior of the stem was almost entirely filled with structureless brown coal, so that only two cross sections could be obtained from the outer parts, one sixteen inches, the other three feet six inches broad. In the first section Goeppert counted 700, in the second 1300 rings of wood, so that for the half-diameter of 5½ feet, at least 2200 rings must have existed. As there is every reason to believe that the rings were formed in earlier ages just as the annual zones are now, this tree would be from 2200 to 2500 years old. Exogenous stems in lignite are often of great size and age. In a trunk near Bonn, Nöggerath counted 792 annual rings. In the turf bogs of the Somme, at Yseux near Abbeville, a trunk of an oak-tree has been found above 14 feet in diameter.
We have thus seen that the vegetation of the globe is represented by numerous distinct floras connected with the different periods of its history, and that the farther back we go, the more are the plants different from those of the present day. There can be no doubt that there have been successive deposits of stratified rocks, and successive creations of living beings. We see that animals and plants have gone through their different phases of existence, and that their remains in all stages of growth and decay have been imbedded in rocks superimposed upon each other in regular succession. It is impossible to conceive that these were the result of changes produced within the limits of a few days. Considering the depth of stratification, and the condition and nature of the living beings found in the strata at various depths, we must conclude (unless our senses are mocked by the phenomena presented to our view) that vast periods have elapsed since the Creator in the beginning created the heavens and the earth. How far it may be possible in the future to correlate the history of the earth inscribed on its rocky tablets and deciphered by the geologist, and that short narrative which forms the introduction[102] to the Sacred Volume, it is too difficult to say. At present there are no satisfactory materials for such a correlation; but one thing is certain, that both Revelation and Geology testify with one voice to the work of a Divine Creator.
"Who shall declare (Hugh Miller remarks) what through long ages the history of creation has been? We see at wide intervals the mere fragments of successive Floras; but know not how, what seem the blank interspaces, were filled; or how, as extinction overtook in succession one tribe of existences after another, and species, like individuals, yielded to the great law of death, yet other species were brought to the birth, and ushered upon the scene, and the chain of being was maintained unbroken. We see only detached bits of that green web which has covered our earth ever since the dry land first appeared. But the web itself seems to have been continuous throughout all time; though, as breadth after breadth issued from the creative loom, the pattern was altered, and the sculpturesque and graceful forms that illustrated its first beginnings and its middle spaces have yielded to flowers of richer colour and blow, and fruits of fairer shade and outline; and for gigantic club-mosses stretching forth their hirsute arms, goodly trees of the Lord have expanded their great boughs; and for the barren fern and the calamite clustering in thickets beside the waters, or spreading on flowerless hill-slopes, luxuriant orchards have yielded their ruddy flush, and rich harvests their golden gleam."
When we find animals and plants, of forms unknown at the present day, in all stages of development, we read a lesson as to the history of the earth's former state as conclusive as that which is derived from the Nineveh relics (independent of Revelation) in regard to the history of the human race. There is no want of harmony between[103] Scripture and Geology. The Word and the Works of God must be in unison, and the more we truly study both, the more they will be found to be in accordance. Any apparent want of correspondence proceeds either from imperfect interpretation of Scripture or from incomplete knowledge of science. The changes in the globe have all preceded man's appearance on the scene. He is the characteristic of the present epoch, and he knows by Revelation that the world is to undergo a further transformation, when the elements shall melt with fervent heat, and when all the present state of things shall be dissolved, ere the ushering in of a new earth, wherein righteousness is to dwell.
Recapitulation of the chief points connected with Fossil Botany:—
1. The vegetation of the globe has varied at different epochs of the earth's history.
2. The farther we recede in geological history from the present day, the greater is the difference between the fossil plants and those which now occupy the surface.
3. All fossil plants may be referred to the great classes of plants of the present day, Acotyledons, Monocotyledons, and Dicotyledons.
4. The fossil species are different from those of the present flora, and it is only when we reach the Tertiary periods that we meet with some genera which are without doubt identical.
5. Fossil plants are preserved in various conditions, according to the nature of their structure, and the mode in which they have been acted upon. Sometimes mere casts of the plants are found, at other times they are carbonised and converted into coal, while at other times, besides being carbonised, they are infiltrated with calcareous or siliceous matter, and finally, they may be petrified.
6. Cellular plants, and the cellular portions of vascular plants, have rarely been preserved, while woody species, and especially Ferns, which are very indestructible, have retained their forms in many instances.
7. In some cases, especially when silicified or charred, the structure[104] of the woody stems can be easily seen in thin sections under the microscope.
8. The determination of fossil plants is a matter of great difficulty, and requires a thorough knowledge of structure, and of the markings on stems, roots, etc.
9. The rocks containing organic remains are called fossiliferous, and are divided into Primary, Secondary, and Tertiary, or into Palæozoic, Mesozoic, and Cainozoic, each of these series being characterised by a peculiar facies of vegetable life.
10. The mere absence of organic remains will not always be a correct guide as to the state of the globe.
11. The number of fossil species has been estimated at between 3000 and 4000; but many parts of plants are described as separate species, and even genera, and hence the number is perhaps greater than it ought to be.
12. Brongniart divides the fossil flora into three great epochs:—1. The reign of Acrogens; 2. The reign of Gymnosperms; 3. The reign of Angiosperms.
13. The reign of Acrogens embraces the Silurian, Carboniferous, and Permian epochs, in which there was a predominance of plants belonging to the natural orders Filices, Lycopodiaceæ, and Equisetaceæ, associated, however, with others of a higher class.
14. The reign of Gymnosperms embraces the lower and middle Secondary periods, and is characterised by the presence of numerous Coniferæ and Cycadaceæ.
15. The reign of Angiosperms includes the Cretaceous and Tertiary periods, and is marked by the predominance of Angiospermous Dicotyledons.
16. Coal is a vague term, referring to all kinds of fuel formed from the chemically-altered remains of plants.
17. When there is a great admixture of mineral matter, so that it will not burn as fuel, then a shale is produced.
18. The microscopic structure of Coal probably varies according to the nature of the plants of which it is composed, and the changes produced by pressure, heat, and other causes. Cellular tissue, punctated woody tissue, and scalariform vessels, have been detected in it.
19. Certain temporary and local floras seem to have given origin to peculiar layers of coal.
20. During the Carboniferous epoch we meet with Ferns, Sigillarias, and their roots called Stigmarias, Lepidodendrons, Ulodendrons, Calamites, Gymnosperms, etc.
21. The plants forming coal have grown in the basin where the coal is found; but sandstone rocks in the coal-measures deposited by water having a considerable velocity, and consequently carrying power, contain sometimes trunks of large trees which have been drifted like snags.
22. The strata between the Permian epoch and Chalk display numerous Gymnosperms, especially belonging to the Cycadaceous Order. Some of them exhibit limited coal deposits.
23. The Chalk and Tertiary strata display not only Acrogens and Gymnosperms, but also Angiospermous Dicotyledons, some of which, at the Miocene period, belong apparently to genera of the present day.
24. Brown Coal occurs in the Upper Tertiary beds, and in it vegetable structure is easily seen under the microscope.
25. Raulin thinks that during the Tertiary epoch the flora of Europe has gradually assumed a more temperate character.
26. The Eocene flora, according to Unger, resembled in many respects that of Australia at the present day.
27. The Miocene flora is characterised by a number of exotic forms of warm regions with those of temperate climates. It is largely seen in the Arctic Regions.
28. The Pliocene flora has great analogy with that of the temperate regions of Europe, North America, and Japan.
On the subject of Fossil Botany the following works may be consulted:—
Abhandlungen der Kaiserlich Königlichen Geologischen Reichsanstalt, Band. ii. Wien. 1855.
Argyll, Duke of, on Tertiary Leaf-Beds in the Isle of Mull, Journ. Geol. Soc. Lond., vii. May 1851.
Balfour, J. H., on certain Vegetable Organisms in Coal from Fordel, Trans. R.S.E., vol. xxi. p. 187.
Baily, W. H., Figures of Characteristic British Fossils, 1871-2.
Bennett, J. Hughes, on the Structure of Torbane Hill Mineral and other Coals, Trans. R. Soc. Ed., vol. xxi. p. 173.
Binney, E. W., on Calamites and Calamodendron, Palæontographical Society's Memoirs, 1868.
——— on the Structure of Fossil Plants found in the Carboniferous Strata. Palæontographical Society's Memoirs, 1871.
——— Description of some Fossil Plants, showing Structure in the[106] Lower Coal Seam of Lancaster and Yorkshire, Phil. Trans., vol. 155, p. 579.
Bowerbank, Fossil Fruits and Seeds of the London Clay.
Brongniart, Histoire des Végétaux Fossiles, 1828-44.
——— Observations sur la Structure intérieure du Sigillaria, etc., in Archives du Museum, i. 405.
——— Exposition Chronologique des Périodes de Végétation, in Ann. des Sc. Nat. 3d series, Bot. xi. 285.
Carruthers, on Gymnospermatous Fruits from the Secondary Rocks of Britain, Journ. Bot., Jan. 1867.
——— on the Structure of the Stems of the Arborescent Lycopodiaceæ of the Coal Measures, Nos. i. to iv., Month. Microsc. Journ., vols. i. ii. iv.
——— on the Cryptogamic Forests of the Coal Period, Paper read before the Royal Institution of Great Britain, 16th April 1869.
——— on the Structure and Affinities of Sigillaria and Allied Genera, Quart. Journ. Geol. Soc., Aug. 1869.
——— on a Fossil Cone from the Coal Measures, Geol. Mag., 1865.
——— on Caulopteris punctata, ibid.
——— on Araucaria Cones from the Secondary Beds of Britain, ibid. 1866.
——— on an Aroideous Fruit from the Stonesfield Slate, ibid. 1867.
——— on Cycadoidea Yatesii, ibid. 1867.
——— on the Structure of the Fruit of Calamites, Journal of Botany, 1867.
——— on British Fossil Pandanaceæ, ibid. 1868.
——— on British Fossil Coniferæ, ibid. 1869.
——— on the Petrified Forest near Cairo, Geol. Mag., vii. 306.
——— on the Structure of a Fern-Stem from the Lower Eocene, Journ. Geol. Soc., xxvi. 349.
——— on the Structure and Affinities of Lepidodendron and Calamites, Trans. Bot. Soc. Edin., viii. 495.
——— on some Fossil Coniferous Fruits, Geol. Mag., vols. iii. vi.
——— on Beania, a new genus of Cycadean Fruit, from the Yorkshire Oolites, Geol. Mag., vol. vi.
——— on Plant-remains from the Brazilian Coal-beds, with Remarks on the genus Flemingites, Geol. Mag., vol. vi.
——— on the Fossil Cycadaceous Stems from the Secondary Rocks of Britain, Linn. Trans., xxvi. 675.
——— on the History and Affinities of the British Coniferæ, Brit. Assoc. Reports, 40th Meeting, p. 71.
Carruthers, List of New Genera and Species of Fossil Plants, Nos. i. ii. and iii., Journal of Botany, vols. viii. ix. x.
Coalfields, by a Traveller under ground.
Corda, Beiträge zur Flora der Vorwelt, Prag. 1845.
Cotta, Dendrolithen, Leipzig, 1850.
Dawson, J. W., on Spore-Cases in Coal, Ann. Nat. Hist., 1871, p. 321.
——— on Vegetable Structures in Coal, Quart. Journ. Geol. Soc., 1860.
——— on the Pre-Carboniferous Flora of New Brunswick and Eastern Canada, Canadian Naturalist, May 1861.
——— on the Flora of the Devonian Period in North-Eastern America, Quart. Journ. Geol. Soc., Nov. 1862.
——— on an Erect Sigillaria and a Carpolite from Nova Scotia, Quart. Journ. Geol. Soc. Lond.
——— on Calamites, Ann. Nat. Hist. 4th ser. vol. iv. 272.
——— on the Varieties and Mode of Preservation of the Fossils known as Sternbergiæ, Canadian Naturalist; also in Edin. New Phil. Journal, N.S. vii. 140.
——— Acadian Geology, 1868.
——— the Fossil Plants of the Devonian and Upper Silurian Formations of Canada, Geol. Survey of Canada, 1871.
——— on the Pre-Carboniferous Floras of North-Eastern America, with special reference to that of the Erian (Devonian) Period, Proc. Roy. Soc. Lond., May 5, 1870.
——— on the Graphite of the Laurentian Rocks of Canada, Quart. Journ. Geol. Soc., xxvi. 112.
Dunker, Zettel, and Meyer, Beiträge zur Naturgeschichte der Vorwelt.
Ettingshausen, Beiträge zur Flora der Vorwelt in Abhandlungen der Geolog. Reichsanstalt, Vienna, 1851.
Forbes, on Tertiary Leaf-Beds in the Isle of Mull, discovered by the Duke of Argyll, F.G.S., with a note on the Vegetable Remains from Ardtun Head, Quart. Journ. Geol. Soc. Lond., vol. vii.
Giebel, Palæontologie.
Goeppert, Beiträge zur Bernsteinflora; sur la Structure de la Houille.
——— Die Gattungen der Fossilen Pflanzen, Bonn, 1841.
——— Monographie des Fossilen Coniferen, 1850.
——— Systema Filicum Fossilium, Nova Acta, xvii.
——— Ueber die Fossilen Cycadeen, Breslau, 1844.
——— Erläuterung der Steinkohlen-Formation.
Goeppert, Die Fossile Flora der Permischen Formation, Palæontographica, Hermann von Meyer, Cassel, 1864.
——— Beiträge zur Kenntniss Fossilen Cycadeen, Breslau.
Grand d'Eury, on Calamites and Asterophyllites, Ann. Nat. Hist., ser. 4, vol. iv. 124.
Harkness, on Coal, Edin. Phil. Journ., July 1854.
Heer, Flora Tertiaria Helvetiæ, 3 vols.
——— Flora Fossilis Arctica, 1868-1871.
——— on the Fossil Flora of Bovey Tracey, Phil. Trans. R.S.L., 152, p. 1039.
——— on the Fossil Flora of North Greenland, Phil. Trans., vol. 159, p. 445.
Hooker, on Some Minute Seed-Vessels (Carpolithes ovulum, Brongniart) from the Eocene beds of Lewisham, Proceed. Geol. Soc., 1855.
——— Vegetation of the Carboniferous Period, in Mem. of Geol. Survey, ii.
——— on a New Species of Volkmannia, Quart. Journ. Geol. Soc. Lond., May 1854.
King, on Sigillaria, etc., in Edin. New Phil. Journal, xxxvi.
Lesquereux, on the Coal Measures of America, Silliman's Journal, 1863.
Lindley and Hutton, Fossil Flora, 3 vols. A revision of the original work, with a supplementary volume containing the recent additions, and a Synopsis of the Fossil Plants of Britain by Mr. W. Carruthers, is announced as about to be published.
Lowry, Table of the Characteristic Fossils of Different Formations.
M'Nab, on the Structure of a Lignite (Palæopitys) from the Old Red Sandstone, Trans. Bot. Soc. Edin., x. 312.
Mueller and Smyth, on Some Vegetable Fossils from Victoria, Geol. Mag., vii. 390.
Meyer, Hermann Von, Palæontographica. Beiträge zur Naturgeschichte der Vorwelt, 1864.
Nicholson, on the Occurrence of Plants in the Skiddaw Slates, Geol. Mag., vol. vi.
Paterson, Description of Pothocites Grantoni, a New Fossil Vegetable from the Coal Formation, Trans. Bot. Soc. Edin., vol. i.
Penny Cyclopædia, vol. vii., Coal Plants.
Pictet, Traité de Paléontologie.
Quekett, on the Minute Structure of Torbane Hill Mineral, Journ. Microsc. Sc., 1854.
Raulin, Flore de l'Europe pendant la Période Tertiaire, in Ann. des Sc. Nat., 3d ser. x. 193.
Redfern, on the Nature of the Torbane Hill and other Varieties of Coal, Brit. Assoc. Liverpool, 1854.
Roehl, A. von, Fossile Flore der Steinkohlen Formation Westphalens.
Saporta, Etudes sur la Végétation du Sud-Est de la France à l'Epoque Tertiaire, Annales des Sciences Naturelles, ser. 4, tome xvi. 309, xvii. 191, xix. 5; ser. 5, tome iii. 5, iv. 5.
Schenk, Professor, die Fossile Flore der Nordwest Deutschen Wealden Formation.
Schimper, Traité de Paléontologie Végétale, 1870-71.
Tate, on the Fossil Flora of the Mountain Limestone Formation of the Eastern Borders, in connection with the Natural History of Coal (in Johnstone's Eastern Borders).
Torbane Coal, as noticed in the Report of the Trial as to the substance called Torbane Mineral or Torbanite.
Unger, Genera et Species Plantarum Fossilium.
——— Chloris Protogæa.
——— Le Monde Primitive (a work which contains picturesque views of the supposed state of the earth at different geological epochs).
——— on the Flora of the Eocene Epoch, Journ. Bot., iii. 39.
Weber and Wersel, Die Tertiarflore der Nieder Sheinescher Braunkohlen Formation.
Williamson, W. C., on the Organisation of the Fossil Plants of the Coal Measures, Ann. Nat. Hist., 1871, p. 134.
——— on the Structure and Affinities of the Plants hitherto known as Sternbergiæ, Mem. Manch. Lit. and Phil. Soc., ix.
——— on a New Form of Calamitean Strobilus, from the Lancashire Coal Measures, Mem. Lit. Phil. Soc. Manchester, vol. iv. 3d series.
——— on the Structure of the Woody Zone of an Undescribed Form of Calamite, Mem. Lit. Phil. Soc. Manchester, vols. iv. and viii. 3d series.
——— on Volkmannia Dawsoni, ibid. 1870-71.
——— on Zamia gigas (Williamsonia gigas), Linn. Trans., xxvi. 663.
——— on the Organisation of Fossil Plants of the Coal Measures, Part I., Calamites, Phil. Trans. R.S.L., vol. 161, p. 477.
Witham, on the Structure of Fossil Vegetables.
Yates, on Zamia gigas, Proceed. Yorkshire Phil. Soc., April 1847.
Young, J., and Armstrong, Jas., on the Carboniferous Fossils of the West of Scotland, Trans. Geol. Soc. Glas., vol. iii.
Besides geological treatises such as those of Ansted,
Beudant, Jukes, Lyell, and others.
PLATE I.
Fig. 1. Palæopteris Hibernica, Schimper (Cyclopteris Hibernica, Forbes). One-sixth the natural size.
Fig. 2. A pinnule somewhat magnified, showing the venation.
Fig. 3. A fertile pinna, natural size.
Fig. 4. Two cup-shaped indusia borne on the rachis.
Fig. 5. Sporangia enclosing spores. From the Coal-measures.
Fig. 6. Sporangia of Hymenophyllum Tunbridgense, Sm. (Fern of present epoch.)
Fig. 7. Sporangium of Polypodium vulgare, Linn. (Fern of present epoch.) Figs. 5, 6, and 7, magnified to the same extent.
Fig. 8. Transverse section of Osmundites Dowkeri, Carruthers.
Fig. 9. Two cells of Osmundites, filled, the one with starch granules, and the other with mycelium of a fungus.
PLATE II.
Fig. 1. Cycadeostrobus ovatus, Carr. From the Wealden, Isle of Wight.
Fig. 2. Beania gracilis, Carr. From the Yorkshire Oolite.
Fig. 3. Bennettites Saxbyanus, Carr. From the Lower Greensand of the Isle of Wight.
Fig. 4. Pinites Leckenbyi, Carr. From the Lower Greensand of the Isle of Wight.
Fig. 5. Trigonocarpon olivæforme, Lindl. and Hutt. From the Coal-measures, Manchester.
Fig. 6. Trigonocarpon sulcatum, Carr. Coal-measures, Wardie, Edinburgh.
Fig. 7. Sequoiites Gardneri, Carr. From the Gault at Folkestone.
Figs. 8, 9. Cupressinites Thujoides, Bowerbank. From the Eocene at Sheppey.
Fig. 10. Scale of Araucarites Brodiei, Carr. From the Great Oolite at Stonesfield.
Fig. 11. Scale of Araucarites Phillipsii, Carr. From the Oolite of Yorkshire.
All the figures on this Plate (except Fig. 2, which is one-half
of the natural size) are drawn the size of nature.
PLATE III.
Fig. 1. Mass of coal from Fordel, Fifeshire, containing numerous sporangia of Flemingites. These sporangia occur in coal from different localities in England and Scotland. Binney has seen them in Wigan coal. Huxley has found them abounding in coal near Bradford (Balfour, R.S.E. Trans.)
Fig. 2. One of the Sporangia entire, and separated from the coal (Balfour).
Fig. 3. Sporangium with its valves separated, containing a quantity of black carbonaceous matter in its interior (Balfour). This matter is formed by the altered spores (microspores).
Fig. 4. Sporangium, showing the triradiate marking on the under surface, and a granulation produced probably by the spores in the interior.
Fig. 5. Punctated woody tissue (Coniferous). From the needle coal of Toplitz, Bohemia (Harkness).
Fig. 6. Scalariform vessels from coal (Balfour).
Fig. 7. Stigmaria, with markings of rootlets. One showing the papilla to which the rootlet was articulated (Hooker).
Fig. 8. Transverse section of Stigmaria, showing the vascular cylinder divided into wedges (Hooker).
Fig. 9. Tissues of Stigmaria, showing the inner portion of the vascular cylinder (Hooker).
Fig. 10. Transverse section of a Lepidostrobus, the fructification of Lepidodendron, showing scales and sporangia (Hooker).
Fig. 11. Ulodendron Taylori (Carruthers).
PLATE IV.
Fig. 1. Sigillaria Brownii, restored (Dawson).
Fig. 2. Sigillaria elegans, restored (Dawson).
Fig. 3. Lepidodendron, restored (Carruthers, Bot. Soc. Trans.)
Fig. 4. Calamites, restored (Carruthers, Bot. Soc. Trans.)
Fig. 5. Psilophyton, a fossil of the Devonian epoch (Dawson).
Abietites, 84, 85, 87.
Acacia, 90, 92.
Acanthocarpum, 72.
Acer, 92, 97.
Acerites, 87.
Acrogens of present day, 26.
Acrogens, fossil, reign of, 25, 26.
Adiantites, 41.
Æthophyllum, 79.
Alder, 94.
Alethopteris, 43, 72.
Algæ, 35.
Algæ of Cretaceous epoch, 87.
Alnites, 87, 94.
Alnus, 94, 97.
Alsophila, 29.
Amber, 90.
Amber flora, Goeppert on the, 91.
Amentiferæ, fossil, 92.
Ancestrophyllum, 48.
Andromeda, 96, 97.
Angiosperms, fossil, reign of, 25, 87.
Annularia, 61, 71.
Anomopteris, 79.
Anona, 97.
Anthodiopsis, 72.
Antholithes, 64.
Anthracite, 36.
Apocynaceæ, fossil, 92.
Araucaria, 5, 6, 7, 85, 90.
Araucarioxylon, structure of, 63.
Araucarites, 82, 83, 84, 85, 86, 87.
Arctic fossil flora (Miocene), 97.
Arctic Regions, Palæozoic flora of, 40.
Arctic Regions, fossil wood of, 95.
Arthropitys, 72.
Artisia, 64.
Asplenium, 28.
Asterophyllites, 35, 61, 71.
Bambusium, 92.
Bauhinia, 90, 92.
Beania, 82.
Bear Island, fossil flora of, 40, 59.
Beeches, 97.
Bennettiteæ, 86.
Bennettites, 85, 87.
Betula, 94, 97.
Bothrodendron, 57.
Bovey Tracey flora, 96.
Bovey Tracey, Devonshire, lignite beds of, 96.
Brachyphyllum, 80, 86, 87.
Bryson's instrument for slitting, 14.
Bucklandia, 84, 86.
Cæsalpinia, 90.
Cainozoic period, fossil plants of, 87.
Calamites, 35, 41, 53.
Calamites, foliage and fruit (woodcut), 62.
Calamites, structure of, 57.
Calamites, structure of fruit, 60.
Calamodendron, 59, 72.
Callipteris, 72.
Callitris, 90.
Camptopteris, 79, 80.
Carboniferous epoch, 36.
Carboniferous vegetation, its general character, 69.
Carbonisation, 9.
[114]Cardiocarpum, 41, 72, 78.
Cardiocarpum, structure of, 64.
Cardiopteris, 40.
Carpinites, 87.
Carpinus, 94.
Carpolithes, 78, 83, 92, 97.
Cassia, 90.
Casts of plants, 8.
Casuarina, 90.
Caulinites, 90.
Caulopteris, 43.
Centrolobium, 90.
Chalk flora, characteristics of, 87.
Chara, 92.
Characeæ, fossil, 91.
Chondrites, 87.
Cinchonaceæ, fossil, 92.
Cinnamomum, 96.
Classes to which fossil plants belong, 2.
Climate as determined by fossil plants, 19.
Climate of the Tertiary period, 100.
Club-mosses, 26, 30.
Coal-basins, 37.
Coal, brown, structure of, 100.
Coal, Fordel, 36, 56.
Coal-formation, extent of, 38.
Coal, household, 36.
Coal-measures, flora of, 39.
Coal, parrot, 36.
Coal-plants, in situ, or drifted, 67.
Coal, structure in, 36.
Coal, Wigan cannel, 36.
Coal of Oolitic epoch, 82.
Coal of Tertiary beds, 100.
Combretaceæ, fossil, 92.
Comptonia, 92, 94.
Comptonites, 87.
Cones, fossil, of Wealden, 85.
Confervites, 87.
Coniferæ, 87.
Coniferæ, modern, 72.
Coniferæ, number of Miocene species, 97.
Coniferæ, Oolitic, 80.
Coniferæ, structure of recent, 74.
Coniferæ of brown coal deposits, 100.
Coniferæ of Miocene Arctic fossil flora, 97.
Coniferæ of Secondary strata, 85.
Coniferæ of Tertiary period, 89.
Coniferous genera of Lias, 79.
Coniferous vegetation of Upper Cretaceous period, appearance of, 89.
Copaifera, 90.
Cordaites, 35, 72.
Cornus, 97.
Corylus, 96, 97.
Cratægus, 97.
Credneria, 87.
Crematopteris, 79.
Cretaceous system, fossil plants of, 87.
Crossozamia, 86.
Cryptogamia, number of Miocene species of, 97.
Cryptomeria, 87.
Cryptomerites, 86.
Ctenis, 78, 79.
Cucumites, 90.
Cunninghamites, 87.
Cupressineæ, 89.
Cupressoxylon, 93.
Cyathea, 29.
Cyatheites, 72.
Cycadaceæ, 87.
Cycadaceæ, fossil, Carruthers' arrangement of, 86.
Cycadaceæ, modern, 72, 75.
Cycadaceæ, Oolitic, 80.
Cycadaceæ in Mesozoic period, 77.
Cycadaceæ of Lias, 79.
Cycadaceæ of Tertiary period, 89.
Cycadaceæ of Wealden epoch, 84.
Cycadeostrobus, 85.
Cycadites, 44, 79, 84, 87.
Cycadoidea, 83.
Cycas, 76.
Cyclopteris, 32, 35, 43, 72.
Cyclostigma, 41.
Cyperites, 48.
Cystoseirites, 87.
Dadoxylon, 35, 63.
Dalbergia, 90.
Dammarites, 87.
Daphnogene, 92, 96, 97.
Dawson on Devonian fossils, 35.
[115]Desmodophyllum, 92.
Dicotyledons of Pliocene epoch, 98.
Dictyothalamus, 72.
Didymophyllum, 48.
Didymosorus, 95.
Dioonopteris, 72.
Dirt-bed, Portland, 83.
Dolichites, 92.
Drepanocarpus, 90.
Dryandroides, 96.
Echitonium, 92, 96.
Encephalartos, 76.
Entada, 90.
Eocene epoch, Algæ of, 90.
Eocene epoch, characteristics of, 90.
Eocene epoch, Coniferæ of, 90, 91.
Eocene epoch, flora of, 89, 90.
Eocene epoch, fruits of, 90.
Eozoon Canadense, 31.
Equisetaceæ, 29, 59.
Equisetites, 71.
Equisetum, 31, 53, 79, 94, 95.
Equisetum spores, 32.
Equisetum, structure of fruit, 60.
Erian fossil plants, 35.
Erythrina, 92.
Exogenous trees of Carboniferous epoch, 62.
Fagus, 94, 97.
Favularia, 46.
Fern-flora in connection with climate, 41.
Ferns, 96.
Ferns, structure of, 29.
Ferns of Carboniferous strata, 41.
Ferns of present day, 26.
Ficus, 96.
Fig, evergreen, 96.
Filicites, 94.
Fittonia, 86, 87.
Flabellaria, 64, 87.
Flemingites, 51, 52, 57.
Floras of present day in connection with fossil plants, 19.
Folliculites, 92.
Fossil botany, recapitulation of chief points connected with, 103.
Fossil botany, list of works treating of, 105.
Fossil plants compared with modern plants, 3, 4.
Fossil plants, determination of, 3.
Fossil plants, mode of preservation of, 8.
Fossil plants, number of, 23.
Fossiliferous periods, according to Brongniart, 25.
Fossiliferous rocks, 20.
Fructification in ferns of Carboniferous epoch, 40.
Fruits, fossil, of Isle of Sheppey, 90.
Fungi, fossil, 91.
Gardenia, 97.
Gault, Coniferæ of, 80, 85.
Getonia, 92.
Glyptostrobus, 97.
Grès bigarré, 78.
Gymnosperms, fossil, reign of, 25.
Gyrogonites, 92.
Haidingera, 78.
Hakea, 97.
Halonia, 57.
Heer's list of plants from the Bovey Tracey Miocene formation, 96.
Heer on the migration of plants, 98.
Heer on the number of species in the Arctic fossil flora, 97.
Heer's remarks on the Polar flora, 98.
Hightea, 90.
Horse-tails, 29.
Huttonia, 71.
Hymenophylleæ, 34.
Hymenophyllites, 71.
Hymenophyllum, 35.
Ilex, 97.
Infiltration, 9.
Inga, 90.
Isoetes, 27, 49, 89.
Ivy, 97.
Juglandites, 87.
Jurassic period of Brongniart, 79.
Kaidacarpum, 84.
[116]Keupric period, 79.
Kimmeridge Clay, Coniferæ of, 85.
Knorria, 41, 48, 57.
Knorria, phyllotaxis of, 55.
Lastrea, 29.
Lauraceæ, 97.
Lauraceæ, fossil, 92.
Laurel, 96.
Laurentian rocks, 31.
Laurus, 92, 96.
Leaf-beds of Ardtun, Mull, 93.
Leaf-beds of Bournemouth, 95.
Leaf-beds, genera of, 94.
Leguminosæ, fossil, 92.
Leguminosites, 90.
Lepidodendron, 35, 41, 49.
Lepidodendron, phyllotaxis of, 54.
Lepidophloios, 57.
Lepidophyllum, 41, 56.
Lepidostrobus, 35, 50, 52.
Lias, Coniferæ of, 80.
Lias, fossil plants of, 79.
Libocedrus, 90.
Lignite, 32.
Lignite beds of Bovey Tracey, 96.
Lignites, 9.
Lonchopteris, 43, 84.
Lough Neagh, Miocene formation of, 93.
Lower Greensand, cone of, 89.
Lower Greensand, Coniferæ of, 85.
Lycopodiaceæ, 49, 54.
Lycopodiaceæ, modern, 26.
Lycopodites, 56.
Lycopodium, 30, 53.
MacClintockia, 97.
Macrospores, 30.
Magnolia, 97.
Mantellia, 83, 84, 86.
Marsilea, 31, 33.
Marsileaceæ, 31.
Mesozoic period, flora of the, 72.
Microspores, 30.
Microzamia, 87.
Mimosa, 90.
Mimosites, 92.
Miocene epoch, flora of, 89, 92.
Miocene period, temperature of, 97.
Mull, leaf-beds of, 93.
Mull, Miocene formation of, 93.
Munsteria, 87.
Myrica, 94, 97.
Naiadaceæ, 87.
Natural orders to which fossil plants belong, 22.
Neuropterideæ, 41.
Neuropteris, 42, 71.
Nicolia, 11.
Nicol's mode of preparing sections, 13.
Nilssonia, 79.
Nipadites, 90.
Noeggerathia, 64, 71, 72.
Nymphæa, 97.
Odontopteris, 42, 72.
Oolitic epoch, flora of, 80.
Oolite, fruits of, 83.
Oolite, Inferior, Coniferæ of, 86.
Oolite, Lower, 82.
Oolite, Scottish, plants of, 81.
Oolite, Upper, 82.
Oolite, Yorkshire, 83.
Osmunda, 89.
Osmundites, 91.
Otopteryx, 97.
Otozamites, 79.
Oxford Clay, Coniferæ of, 86.
Palæophytology, 1.
Palæopitys, 32.
Palæopteris, 32, 34, 41.
Palæozamia, 79.
Palæozoic or Primary period, 26.
Palæozoology, 1.
Palissya, 80, 86.
Paliurus, 97.
Palm, 96.
Palmacites, 87, 90, 96.
Pandanaceæ, 84.
Pecopteris, 42, 96.
Pecopterideæ, 41.
Pepperworts, 31.
Permian flora, 71.
Permian period, fruits of, 72.
Petrifaction, 9.
Petrified forests, 11.
Pence, 64, 80, 82, 86, 89.
[117]Phanerogamia, number of Miocene species of, 97.
Phaseolites, 92.
Phœnicites, 92.
Phyllotaxis, 54, 55.
Pilularia, 31.
Pinites, 78, 85, 86, 87, 89, 100.
Pinites, structure of, 63.
Pinus, 86, 94, 97.
Pissadendron, 63.
Pitus, structure of, 64.
Plane, 94.
Platanites, 94.
Platanus, 97.
Pliocene epoch, flora of the, 89, 98.
Plumiera, 92.
Podocarpus, 90.
Podocarya, 84.
Portland beds, 82.
Portland Crag, 82.
Portland stone, Coniferæ of, 85.
Pothocites, 66.
Proteaceæ, fossil, 92, 97.
Protopteris, 87.
Prototaxites, 35.
Prunus, 97.
Psaronius, 44, 71.
Psilophyton, 35.
Pterocarpus, 90.
Pterophyllum, 84, 79.
Purbeck, Coniferæ of, 85.
Purbeck period, 83.
Quercus, 94, 96, 97.
Raulin on the Tertiary flora of Central Europe, 99.
Raumeria, 86.
Recapitulation of chief points connected with fossil botany, 103.
Rhabdocarpum, 72, 77.
Rhamnites, 94, 95.
Rhamnus, 94, 97.
Rhizocarpeæ, 31.
Rings, number of annual, in fossil Exogens, 100.
Sagenopteris, 71, 79.
Salicites, 87.
Salix, 97.
Sargassites, 87.
Scalariform vessels, 30.
Schizopteris, 43.
Secondary period, flora of the, 72.
Sections of fossils for microscope, 12.
Selaginella, 27, 51, 53.
Selaginites, 35.
Senftenbergia, 40.
Sequoia, 87, 96, 97.
Sequoiites, 85, 89.
Shale, 37.
Sheppey, fruits of Isle of, 90.
Sigillaria, 45.
Silicified stems, 10.
Sphenophyllum, 35, 61.
Sphenopterideæ, 41.
Sphenopteris, 34, 41, 42.
Sporangia, 30, 56.
Stangeria, 78.
Steinhauera, 92.
Sternbergia, 64, 97.
Stigmaria, 41, 47, 48.
Stonesfield slate, 82.
Stratified rocks, 21.
Structure of fossil plants, 12.
Table of formations, 21.
Taxites, 86, 94, 95, 100.
Taxodieæ, 89.
Taxodites, 79, 80, 94, 95.
Taxodium, 97.
Terminalia, 92.
Tertiary flora of Europe, 99.
Tertiary period, characteristics of, 89, 100.
Tertiary period, fossil plants of, 87.
Tertiary vegetation, Brongniart's divisions of, 89.
Thaumatopteris, 80.
Thuites, 81, 85, 86.
Thujopsis, 97.
Tilia, 97.
Trap rocks, 20.
Tree-fern, 27.
Trees of Miocene Arctic fossil flora, 97.
Triassic fossils, 77.
Trigonocarpum, 64, 72.
Triplosporites, 50, 53.
Tuff-beds, 94.
[118]Tulip tree, 97.
Ulmus, 92.
Ulodendron, 57.
Underclay, 37.
Unger's list of genera of Eocene epoch, 90.
Upper Chalk, 85.
Upper Greensand, Coniferæ of, 85.
Vaccinium, 96.
Vitis, 96.
Volkmannia, 60.
Voltzia, 78, 79.
Vosgesian period, Brongniart's, 78.
Walchia, 71.
Walnuts, 97.
Wealden, Coniferæ of, 85.
Wealden epoch, flora of, 84.
Widdringtonites, 87.
Williamsonia, 81.
Williamsonieæ, 86.
Willow, 97.
Works, list of, treating of fossil botany, 105.
Yatesia, 86.
Yew, 94.
Zamia, 78.
Zamieæ, 86.
Zamiostrobus, 78.
Zamites, 78, 79, 84, 87.
Zostera, 32.
Zosterites, 87.
THE END.
Printed by R. & R. Clark, Edinburgh.
[1] Miller's Footprints of the Creator, 192-199. Doubts have been thrown on the antiquity of this specimen by those who support the erroneous progressive development theory; but the presence, in the same nodule, of a scale of a fish only found in the lower Old Red, puts the matter beyond doubt. Dr. M'Nab on the Structure of a Lignite (Palæopitys) from the Old Red Sandstone. (Trans. Bot. Soc. x. p. 312.)
[2] Specimens of these fossil plants, as well as numerous others, illustrating the fossil flora of Scotland, are to be seen in Mr. Miller's collection, now in the Edinburgh Museum of Science and Art.
[3] Dawson, Jour. Geol. Soc. Lond. xv. Canadian Naturalist, v. Acadian Geology, 2d edit. Fossil plants of the Devonian and upper Silurian Formations of Canada, with 20 plates; in Report of Geological Survey of Canada.
[4] Maclaren, Geology of Fife and the Lothians, p. 116.
[5] Our Coal-fields, by a Traveller under Ground.
[6] See Hall's Coal-fields of Great Britain, 1861; Roscoe's Lectures on Coal, Manchester, 1866-67; Hunt's Mineral Statistics of Great Britain; Taylor's Statistics of Coal, 1855-56.
[7] Heer, Flora fossilis Arctica; Fossile Flora der Bären Insel., 1871.
[8] In giving names to fossil Ferns, the Greek word πτερίς, meaning a Fern, is often used with a prefix indicating some character in the form of the leaves, or stem, or fructification: such as, πέκος, a comb; νεῦρον, a nerve; ὀδούς, a tooth; σφήν, a wedge; καυλός, a stalk or stem; κύκλος, a circle; σχίζω, a split, etc.
[9] The imbedding of plants in an erect state in strata is similar to what was noticed at the present day by Gardner in Brazil, where stems of recent Coco-nut Palms were seen covered with sand to the depth of 50 feet.
[10] For woodcuts 44, 47, and 48, I am indebted to Dr. H. Bence Jones, who has kindly placed them at my disposal. They were used to illustrate Mr. Carruthers' remarks on the Cryptogamic forests of the Coal period, published in the Journal of the Royal Institution of Great Britain, April 16, 1869. Mr. Carruthers' observations are given in the text.
[11] Conjugate spirals result from whorls of usually 2, 3, 5, 8, etc., leaves arranged so as to give 2, 5, 8, etc., parallel spirals, each with an angular divergence equal to ½, ⅓, ⅕, ⅛, etc., of one of the fractions expressing the divergence in an arrangement of alternate leaves.
[12] By inadvertence, the diameter is stated in my Class-book as 4-5 inches.
[13] See Remarks on the Structure of Calamites by W. C. Williamson, Philos. Trans., 161, p. 477.
[14] Williamson on the Structure and Affinities of Sternbergiæ, in Manch. Lit. and Phil. Soc. Mem. ix. Dawson on Sternbergia, in Edin. New Phil. Journ., new series, vii. 140.
[15] See Notice of Antholithes Pitcairniæ, by C. W. Peach, in Bot. Soc. Trans. Edin. vol. xi.
[16] See Professor Duns on the association of Cardiocarpum with Sphenopteris. Proc. R.S.E., April 1, 1872.
[17] See Meyer's Palæontographica, Cassel, 1864.
[18] See fuller description of Coniferæ and Cycadaceæ in Balfour's Class Book of Botany, pp. 906-912.
[19] Coal in the Kimmeridge clay is probably of animal origin.
[20] Carruthers, Geol. Mag., vol. viii. December 1871.
[21] Annals and Mag. of Nat. Hist. 2d ser. ii. 380.
[22] Journ. Geol. Soc. of London, vii.
[23] Philosophical Transactions, R. Soc. Lond., vol. clii. p. 1039.
[24] Heer, Flore Fossile des Regions Polaires, Zurich; also Bibliotheque Univ. xxxix. p. 12; see also Ann. Nat. Hist. 4th ser. i. 61, iv. 81.
[25] Raulin, Sur les Transformations de la Flore de l'Europe centrale pendant la période Tertiaire.—Ann. des Sc. Nat. 3d ser. Bot. x. 193.
PROFESSOR BALFOUR'S
BOTANY.
In one vol., royal 8vo, pp. 1117, with 1800 Illustrations, price 21s.,
CLASS-BOOK OF BOTANY.
Being an Introduction to the Study of the Vegetable Kingdom.
By J. HUTTON BALFOUR, M.D., F.R.S.,
Professor of Medicine and Botany in the University of Edinburgh, Regius Keeper of the Royal Botanic Garden, and Queen's Botanist for Scotland.
(May also be had in two Parts, price 21s.)
"In Dr. Balfour's 'Class-Book of Botany,' the author seems to have exhausted every attainable source of information. Few, if any, works on this subject contain such a mass of carefully collected and condensed matter, and certainly none are more copiously or better illustrated."—Hooker's Journal of Botany.
"Professor Balfour's 'Class Book of Botany' is too well and favourably known to botanists, whether teachers or learners, to require any introduction to our readers. It is, as far as we know, the only work which a lecturer can take in his hand as a safe text-book for the whole of such a course as is required to prepare students for our University or medical examinations. Every branch of botany, structural and morphological, physiological, systematic, geographical, and palæontological, is treated in so exhaustive a manner, as to leave little to be desired.
"The work is one indispensable to the class-room, and should be in the hands of every teacher."—Nature.
"The voluminous and profusely illustrated work of Dr. Balfour is too well known to need any words of comment."—Lancet.
EDINBURGH: ADAM AND CHARLES BLACK.
JUKES' GEOLOGY.
Just Published, in crown 8vo, cloth, price 12s. 6d.,
NEW EDITION OF BEETE JUKES'
MANUAL OF GEOLOGY.
THIRD EDITION.
Illustrated with numerous Woodcuts.
Edited by ARCHIBALD GEIKIE, F.R.S.,
Professor of Geology and Mineralogy in the University of Edinburgh, and Director of the Geological Survey of Scotland.
"A book which every earnest student of geology will welcome with delight, and than which he can find no better guide to his studies."—Edinburgh Courant.
OWEN'S PALÆONTOLOGY.
Second Edition, 8vo, illustrated, price 7s. 6d.
PALÆONTOLOGY,
OR
A SYSTEMATIC SUMMARY OF EXTINCT ANIMALS AND THEIR GEOLOGICAL RELATIONS.
By RICHARD OWEN, F.R.S.,
Superintendent of the Natural History Department in the British Museum.
"The Prince of Palæontologists has here presented us with a most comprehensive survey of the characters, succession, geological position, and geographical distribution of the various forms of life that have passed away."—Medical Times and Gazette.
EDINBURGH: ADAM AND CHARLES BLACK.
TRANSCRIBER'S NOTE
Basic fractions are displayed as ½ ⅓ ¼ etc; other fractions are shown in the form a/b, eg 3/11 or 13/(34×2).
Most entries in the Table of Contents had a corresponding section heading in the text. Twelve entries had a corresponding page-header, on odd-numbered pages, but no section heading in the text itself. All the page-headers have of course been removed in the etext. To improve readability these twelve section headings have been created and inserted in the etext; they have been italicized to indicate they have been added by the transcriber.
Part of the table at page 22, items 30-38, has been slightly restructured in its bracketing. There is no loss of information.
The caption for an illustration is displayed as a sidenote in the etext. It was shown as a page footnote in the original text.
Obvious typographical errors and punctuation errors have been corrected after careful comparison with other occurrences within the text and consultation of external sources.
Except for those changes noted below, misspelling in the text, and inconsistent or archaic usage, have been retained. For example, planished; punctated; coal-field, coalfield; criddles.
Pg 11, 'silicicified' replaced by 'silicified'.
Pg 39, '1-20th' replaced by '1/20th' for consistency.
Pg 42 Footnote [8], 'I split' replaced by 'a split'.
Pg 73 Illustration, 'Fg. 61' replaced by 'Fig. 61'.
Pg 79, 'aborescent' replaced by 'arborescent'.
Pg 102, 'to difficult' replaced by 'too difficult'.
Pg 105, '29. The Pliocene' replaced by '28. The Pliocene'.
Pg 111, 'Erom the Gault' replaced by 'From the Gault'.
End of the Project Gutenberg EBook of Introduction to the Study of
Palæontological Botany, by John Hutton Balfour
*** END OF THIS PROJECT GUTENBERG EBOOK INTRODUCTION--PALAEONTOLOGICAL BOTANY ***
***** This file should be named 50882-h.htm or 50882-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/5/0/8/8/50882/
Produced by Brian Coe, John Campbell and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive/American Libraries.)
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.